

The ring-infected erythrocyte surface antigen (RESA) of Plasmodium falciparum stabilizes spectrin tetramers and suppresses further invasion
- PMID: 17468340
- PMCID: PMC1924765
- DOI: 10.1182/blood-2007-02-076919
The ring-infected erythrocyte surface antigen (RESA) of Plasmodium falciparum stabilizes spectrin tetramers and suppresses further invasion
Abstract
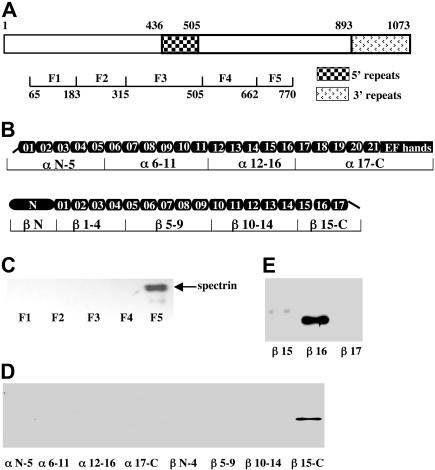
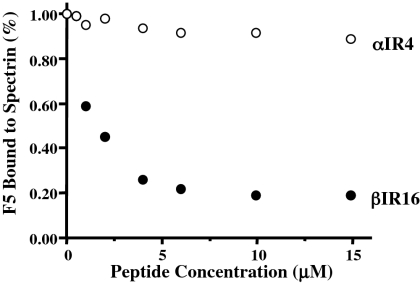
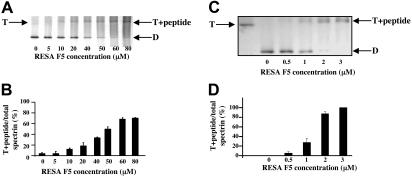
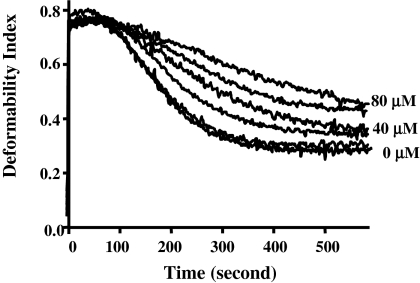
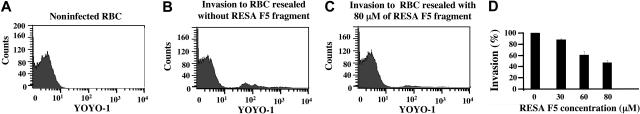
The malaria parasite Plasmodium falciparum releases the ring-infected erythrocyte surface antigen (RESA) inside the red cell on entry. The protein migrates to the host cell membrane, where it binds to spectrin, but neither the nature of the interaction nor its functional consequences have previously been defined. Here, we identify the binding motifs involved in the interaction and describe a possible function. We have found that spectrin binds to a 108-amino acid fragment (residues 663-770) of RESA, and that this RESA fragment binds to repeat 16 of the beta-chain, close to the labile dimer-dimer self-association site. We further show that the RESA fragment stabilizes the spectrin tetramer against dissociation into its constituent dimers, both in situ and in solution. This is accompanied by enhanced resistance of the cell to both mechanical and thermal degradation. Resealed erythrocytes containing RESA(663-770) display resistance to invasion by merozoites of P falciparum. We infer that the evolutionary advantage of RESA to the parasite lies in its ability to prevent invasion of cells that are already host to a developing parasite, as well as possibly to guard the cell against thermal damage at the elevated body temperatures prevailing in febrile crises.
Figures
References
-
- Hiller NL, Bhattacharjee S, van Ooij C, et al. A host-targeting signal in virulence proteins reveals a secretome in malarial infection. Science. 2004;306:1934–1937. - PubMed
-
- Baruch DI, Pasloske BL, Singh HB, et al. Cloning the P. falciparum gene encoding PfEMP1, a malarial variant antigen and adherence receptor on the surface of parasitized human erythrocytes. Cell. 1995;82:77–87. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
