

Review
doi: 10.1016/j.sleep.2007.03.011.
Epub 2007 Apr 30.
Sleep-dependent memory consolidation and reconsolidation
Affiliations
- PMID: 17470412
- PMCID: PMC2680680
- DOI: 10.1016/j.sleep.2007.03.011
Item in Clipboard
Review
Sleep-dependent memory consolidation and reconsolidation
Sleep Med.
2007 Jun.
Abstract
Molecular, cellular, and systems-level processes convert initial, labile memory representations into more permanent ones, available for continued reactivation and recall over extended periods of time. These processes of memory consolidation and reconsolidation are not all-or-none phenomena, but rather a continuing series of biological adjustments that enhance both the efficiency and utility of stored memories over time. In this chapter, we review the role of sleep in supporting these disparate but related processes.
Figures
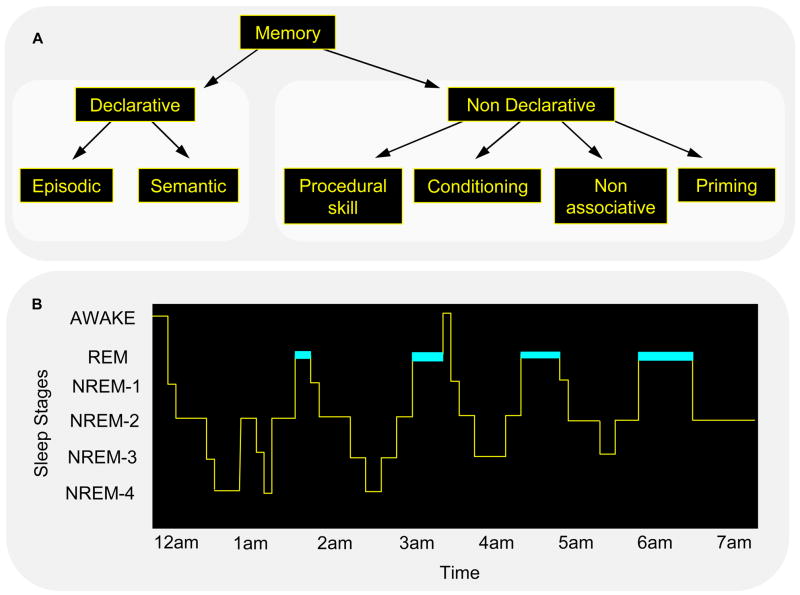
Forms of memory and stages of sleep. Neither memory (A) nor sleep (B) represents a homogeneous phenomenon. (A) Declarative memory includes consciously accessible memories of fact-based information (i.e., knowing “what”), and contains several subcategories, including episodic memory (memory for events in one’s past) and semantic memory (memory for general knowledge) [110]. In contrast, non-declarative memory includes all non-conscious memories, and includes subcategories such as conditioning, implicit memory and procedural memory (i.e., knowing “how”). (B) In mammals, sleep is divided into REM and NREM sleep, and in primates and felines, NREM sleep has been divided into sub-stages 1 through 4, corresponding to increasingly deeper states of sleep [7]. The deepest NREM stages, stages 3 and 4, are collectively referred to as slow wave sleep (SWS), based on a prevalence of low frequency (0.5–4 Hz) cortical oscillations. Dramatic changes in brain electrophysiology, neurochemistry and functional anatomy occur across these sleep stages, making them biologically distinct from the waking brain, and dissociable from one another. For example, SWS is characterized by a diminution in cholinergic activity and REM sleep by a suppression of release of norepinephrine from the locus coeruleus and serotonin from the raphe nucleus. (Reproduced with permission from [8])
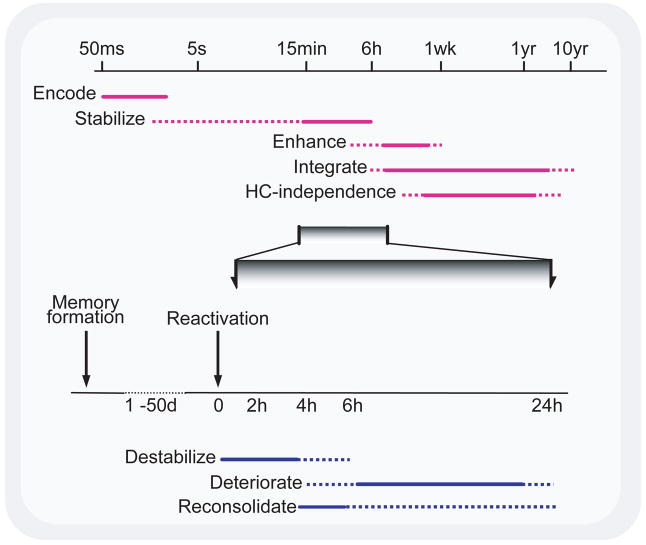
Time course of memory processes. (Top) Memory formation and consolidation – after the initial rapid encoding of a sensory experience, the neural representation of the memory can go through a number of automatic processes, independent of rehearsal, intent, or awareness. These can stabilize and enhance a memory, making it resistant to interference and more effective to guiding behavior, and also integrating the memory into larger associative networks. The latter process is thought to permit episodic memories to be recalled without hippocampal (HC) involvement. The extent to which such processes affect different memory systems is unclear. Note logarithmic time scale. (Bottom) Memory reactivation and reconsolidation – after stabilization is complete, reactivation of a memory can lead to its return to an unstable form. Normally, such memories appear to be reconsolidated following this destabilization, but if such reconsolidation is blocked, degradation of the memory can ensue. (Reproduced with permission from [8])
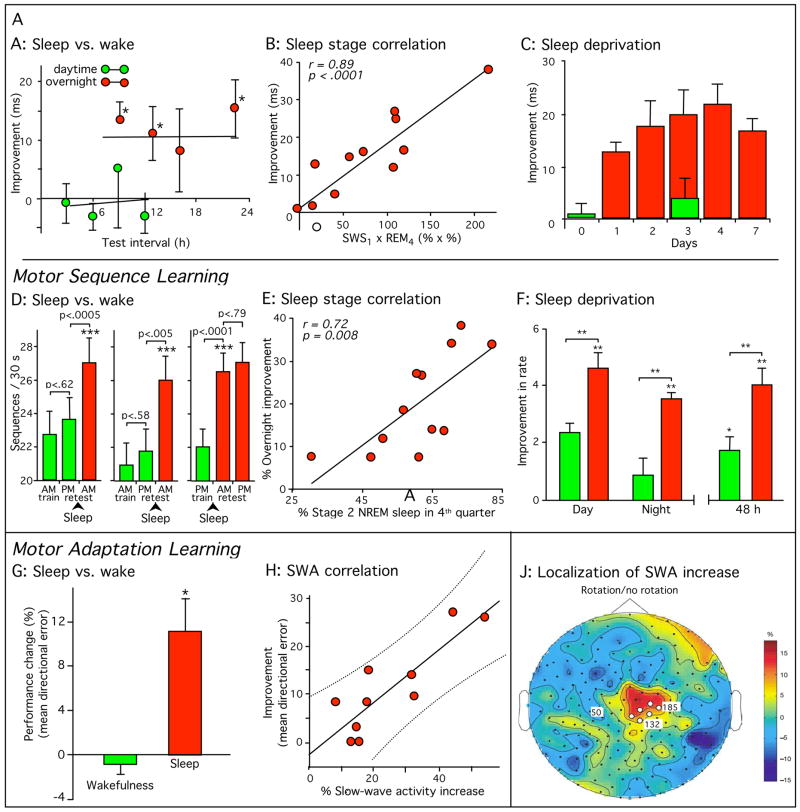
Sleep-dependent consolidation of procedural memories. (A–C) Participants in a visual texture discrimination task show improvement only after post-training sleep. Improvement correlated with early-night slow-wave sleep and late-night REM sleep. (D–F) Participants in a motor sequence finger-tapping task show similar sleep-dependent improvement, even when finger movement was suppressed with mittens during wake periods (D, right panel), improvement which correlated with late-night stage 2 non-REM sleep. (G-J) Participants in a motor adaptation task also show sleep-dependent improvement, correlated with EEG slow-wave activity in task-related regions of the cortex. All error bars represent standard error of the mean. Green bars, performance without intervening sleep or without sleep on the first post-training night. Dark red bars, performance after normal sleep. (Reproduced with permission from [8]; Panel J with permission from [45])

Serial Reaction Time Task. Four circles are presented on a computer screen and lit in a repeated, pseudorandom order. When each circle is lit, the button beneath it must be pressed. In this example, the fourth, then first, then third button are lit sequentially. To determine whether subjects are learning the finger movements (as a motor task) or the spatial positions (as a visual task), subjects are retested using their other hand. For example, subjects trained with their right hand are retested with their left hand. For the spatial sequence 4–1–3 shown here, during training subjects would press keys with the little, then index, and then ring finger of their right hand. If asked to type the same spatial sequence (4–1–3) at retest, subjects would have to use the index, little, and then middle fingers of their left hand. In contrast, to test for retention of learning of the motor sequence (little–index–ring fingers), subjects would be retested on the sequence 1–4–3 in place of 4–1–2.

Sleep-dependent motor memory reorganization in the human brain. Subjects were trained on the sleep-dependent finger-tapping motor skill task and then tested 12 hours later, either following a night of sleep or a day of wake, during a functional magnetic resonance imaging (fMRI) brain-scanning session. Scans after sleep and wake were compared (subtracted), resulting in regions showing increased fMRI activity post-sleep (in red/yellow; A–D) or decreased signal activity (in blue; E–H) post-sleep, relative to post-wake. Activation patterns are displayed on three-dimensional rendered brains (top panel of each graphic), together with corresponding coronal sections (bottom panel of each graphic). Coronal sections all show left on the left; three-dimensional renderings are reversed left for right except (A). Following sleep, regions of increased activation were identified in the left cerebellum (A), the right primary motor cortex (B), the right hippocampus (C), and the right medial prefrontal cortex (D). Regions of decreased activity post-sleep were expressed bilaterally in the parietal lobes (E), together with the left insula cortex (F), left temporal pole (G), and left frontopolar area (H), all regions of the extended limbic system. All data are displayed at a corrected threshold of p < 0.05. (Reproduced with permission from [111].
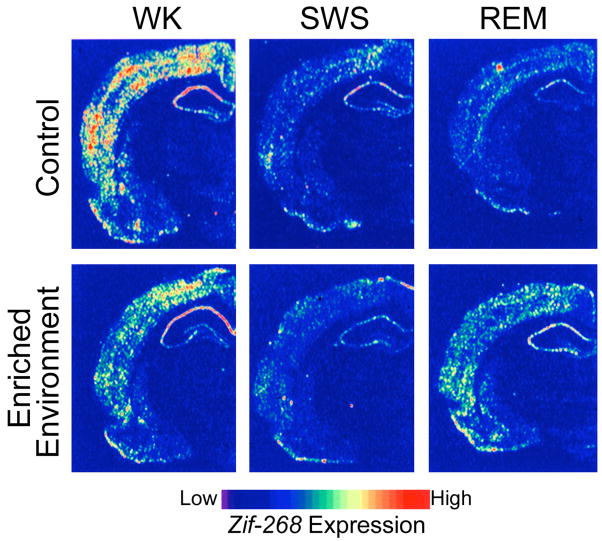
Experience-dependent up-regulation of the synaptic plasticity related immediate early gene zif-268 during periods of wakefulness and REM sleep in the rat. Autoradiograms of frontal coronal brain sections in which gene-expression levels best represent the means for each group studied. (Top panels) In controls, zif-268 expression decreased from wake (WK) to SWS and REM. (Bottom panels) In enriched-environment animals, zif-268 levels decreased from WK to SWS, but then increased in REM. This effect was particularly noticeable in the cerebral cortex and the hippocampus. (Reproduced with permission from [107])
References
-
- Schacter D, Tulving E. What are the memory systems of 1994? In: Tulving E, editor. Memory systems 1994. viii. The MIT Press; 1994. pp. 1–38.
-
- Misanin JR, et al. Retrograde amnesia produced by electroconvulsive shock after reactivation of a consolidated memory trace. Science. 1968;160:554–555. - PubMed
-
- Schneider AM, Sherman W. Amnesia: a function of the temporal relation of footshock to electroconvulsive shock. Science. 1968;159:219–221. - PubMed
-
- McGaugh JL. Memory--a century of consolidation. Science. 2000;287:248–251. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
