

RINT-1 serves as a tumor suppressor and maintains Golgi dynamics and centrosome integrity for cell survival
- PMID: 17470549
- PMCID: PMC1951495
- DOI: 10.1128/MCB.02396-06
RINT-1 serves as a tumor suppressor and maintains Golgi dynamics and centrosome integrity for cell survival
Abstract
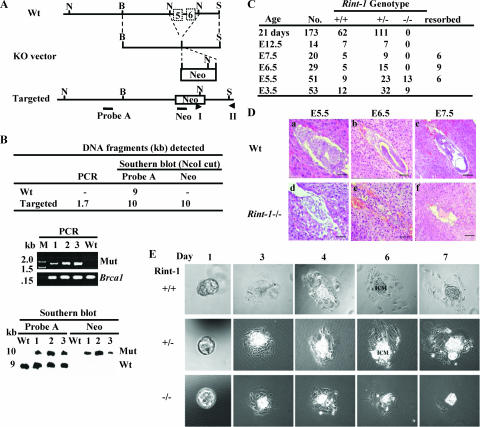
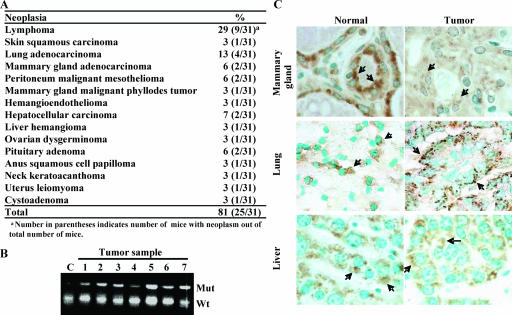
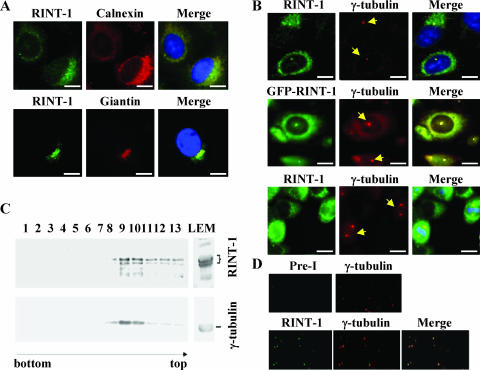
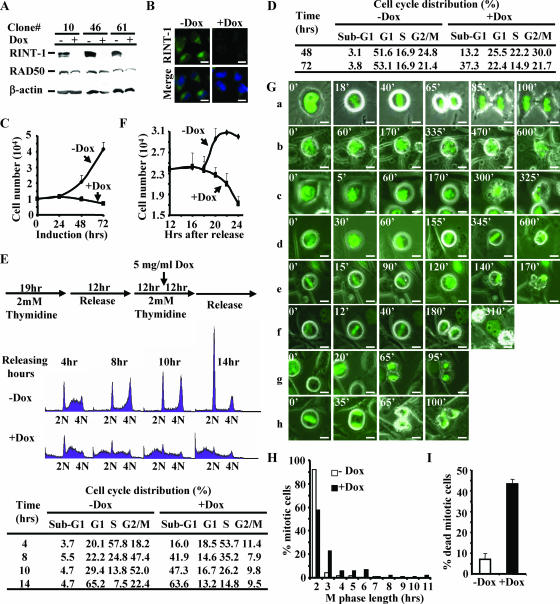
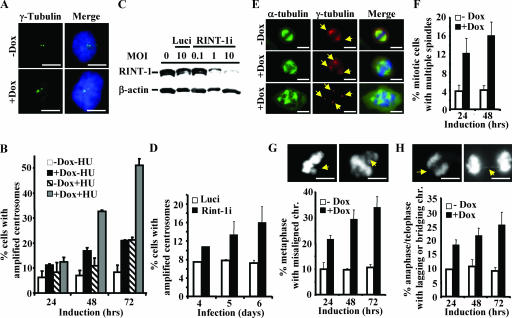
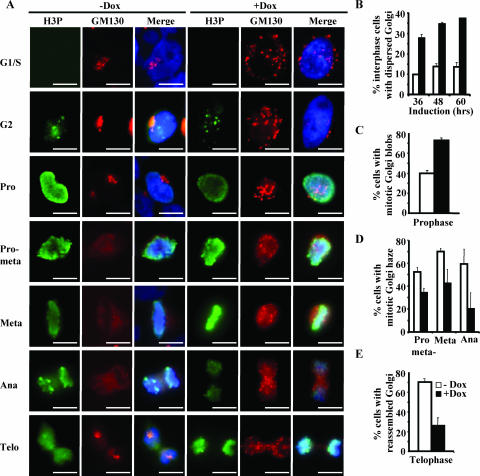
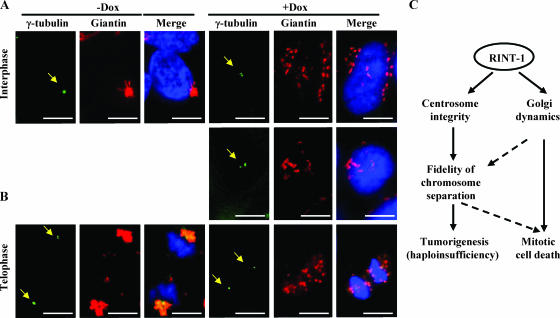
Faithful mitotic partitioning of the Golgi apparatus and the centrosome is critical for proper cell division. Although these two cytoplasmic organelles are probably coordinated during cell division, supporting evidence of this coordination is still largely lacking. Here, we show that the RAD50-interacting protein, RINT-1, is localized at the Golgi apparatus and the centrosome in addition to the endoplasmic reticulum. To examine the biological roles of RINT-1, we found that the homozygous deletion of Rint-1 caused early embryonic lethality at embryonic day 5 (E5) to E6 and the failure of blastocyst outgrowth ex vivo. About 81% of the Rint-1 heterozygotes succumbed to multiple tumor formation with haploinsufficiency during their average life span of 24 months. To pinpoint the cellular function of RINT-1, we found that RINT-1 depletion by RNA interference led to the loss of the pericentriolar positioning and dispersal of the Golgi apparatus and concurrent centrosome amplification during the interphase. Upon mitotic entry, RINT-1-deficient cells exhibited multiple abnormalities, including aberrant Golgi dynamics during early mitosis and defective reassembly at telophase, increased formation of multiple spindle poles, and frequent chromosome missegregation. Mitotic cells often underwent cell death in part due to the overwhelming cellular defects. Taken together, these findings suggest that RINT-1 serves as a novel tumor suppressor essential for maintaining the dynamic integrity of the Golgi apparatus and the centrosome, a prerequisite to their proper coordination during cell division.
Figures
References
-
- Ajiro, K., K. Yoda, K. Utsumi, and Y. Nishikawa. 1996. Alteration of cell cycle-dependent histone phosphorylations by okadaic acid. Induction of mitosis-specific H3 phosphorylation and chromatin condensation in mammalian interphase cells. J. Biol. Chem. 271:13197-13201. - PubMed
-
- Altan-Bonnet, N., R. Sougrat, and J. Lippincott-Schwartz. 2004. Molecular basis for Golgi maintenance and biogenesis. Curr. Opin. Cell Biol. 16:364-372. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous