

Hif-1alpha regulates differentiation of limb bud mesenchyme and joint development
- PMID: 17470636
- PMCID: PMC2064828
- DOI: 10.1083/jcb.200612023
Hif-1alpha regulates differentiation of limb bud mesenchyme and joint development
Abstract
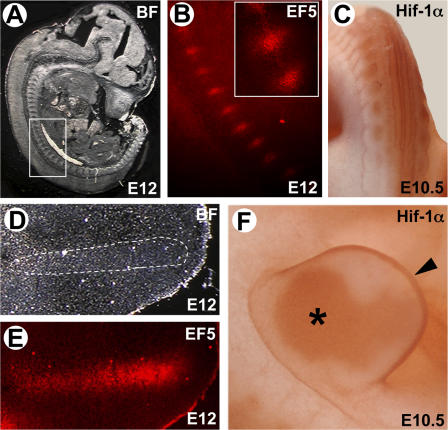
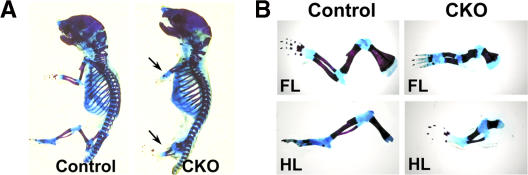
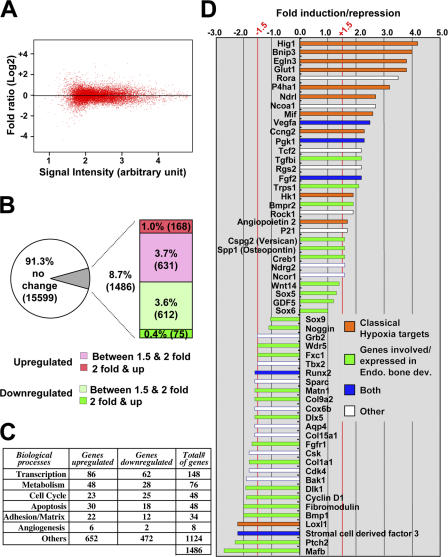
Recent evidence suggests that low oxygen tension (hypoxia) may control fetal development and differentiation. A crucial mediator of the adaptive response of cells to hypoxia is the transcription factor Hif-1alpha. In this study, we provide evidence that mesenchymal condensations that give origin to endochondral bones are hypoxic during fetal development, and we demonstrate that Hif-1alpha is expressed and transcriptionally active in limb bud mesenchyme and in mesenchymal condensations. To investigate the role of Hif-1alpha in mesenchymal condensations and in early chondrogenesis, we conditionally inactivated Hif-1alpha in limb bud mesenchyme using a Prx1 promoter-driven Cre transgenic mouse. Conditional knockout of Hif-1alpha in limb bud mesenchyme does not impair mesenchyme condensation, but alters the formation of the cartilaginous primordia. Late hypertrophic differentiation is also affected as a result of the delay in early chondrogenesis. In addition, mutant mice show a striking impairment of joint development. Our study demonstrates a crucial, and previously unrecognized, role of Hif-1alpha in early chondrogenesis and joint formation.
Figures







References
-
- Bi, W., J.M. Deng, Z. Zhang, R.R. Behringer, and B. de Crombrugghe. 1999. Sox9 is required for cartilage formation. Nat. Genet. 22:85–89. - PubMed
-
- Brunet, L.J., A. McMahon, and R.M. Harland. 1998. Noggin, cartilage morphogenesis, and joint formation in the mammalian skeleton. Science. 280:1455–1457. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
