

NF90 regulates inducible IL-2 gene expression in T cells
- PMID: 17470640
- PMCID: PMC2118567
- DOI: 10.1084/jem.20052078
NF90 regulates inducible IL-2 gene expression in T cells
Abstract
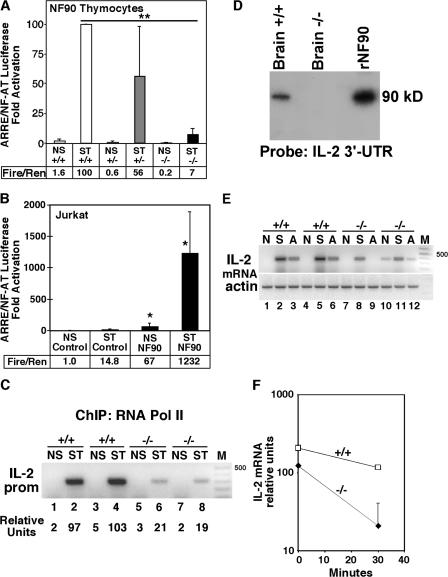
Activation of T cells induces the production of T cell growth and survival factor interleukin (IL) 2. Regulatory T cells intrinsically fail to induce IL-2 expression upon activation and can suppress IL-2 production in conventional T cells. Thus, the control of IL-2 expression is critically important to T cell immune responses, yet the mechanisms remain incompletely understood. Nuclear factor (NF) 90 is a zinc-finger DNA- and double-stranded RNA-binding protein subunit that binds specifically to the antigen receptor response element (ARRE)/NF of activated T cells target sequence in the IL-2 proximal promoter. Inducible binding of NF90 to the IL-2 promoter in vivo is shown by chromatin immunoprecipitation. NF90 gene-targeted mice exhibit perinatal lethality. Compared with newborn NF90(+/+) mice, newborn NF90(-/-) mice demonstrate severe impairment of IL-2 expression. Compared with wild-type cells, T cells deficient in NF90 are impaired in ARRE and IL-2 transcriptional activation and IL-2 mRNA stabilization. Fetal liver cells from NF90 gene-targeted mice were transplanted into irradiated adult recombination activating gene (RAG)-2(-/-) and IL-2Rgamma(-/-) mice deficient in T cells, B cells, and natural killer cells. NF90(+/+)- and NF90(-/-)-RAG chimeric mice showed grossly normal repopulation of the thymus and spleen, but only NF90(-/-) T cells were severely impaired in IL-2 gene expression. Compared with littermates, NF90(-/-) RAG chimeric mice exhibited profound T cell lymphocytopenia in the peripheral circulation. Thus, NF90 regulates inducible IL-2 transcription, mRNA stability, and gene expression in T cells and represents a novel therapeutic target for the modulation of T cell immune responses.
Figures


References
-
- Kao, P.N., L. Chen, G. Brock, J. Ng, J. Kenny, A.J. Smith, and B. Corthesy. 1994. Cloning and expression of cyclosporin A- and FK506-sensitive nuclear factor of activated T-cells: NF45 and NF90. J. Biol. Chem. 269:20691–20699. - PubMed
-
- Corthesy, B., and P.N. Kao. 1994. Purification by DNA affinity chromatography of two polypeptides that contact the NF-AT DNA binding site in the interleukin 2 promoter. J. Biol. Chem. 269:20682–20690. - PubMed
-
- Shim, J., H. Lim, J.R. Yates, and M. Karin. 2002. Nuclear export of NF90 is required for interleukin-2 mRNA stabilization. Mol. Cell. 10:1331–1344. - PubMed
-
- Xu, Y.H., and G.A. Grabowski. 1999. Molecular cloning and characterization of a translational inhibitory protein that binds to coding sequences of human acid beta-glucosidase and other mRNAs. Mol. Genet. Metab. 68:441–454. - PubMed
-
- Shi, L., G. Zhao, D. Qiu, W.R. Godfrey, H. Vogel, T.A. Rando, H. Hu, and P.N. Kao. 2005. NF90 regulates cell cycle exit and terminal myogenic differentiation by direct binding to the 3′-untranslated region of MyoD and p21WAF1/CIP1 mRNAs. J. Biol. Chem. 280:18981–18989. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
