

Class IA phosphoinositide 3-kinases are obligate p85-p110 heterodimers
- PMID: 17470792
- PMCID: PMC1876529
- DOI: 10.1073/pnas.0700373104
Class IA phosphoinositide 3-kinases are obligate p85-p110 heterodimers
Abstract
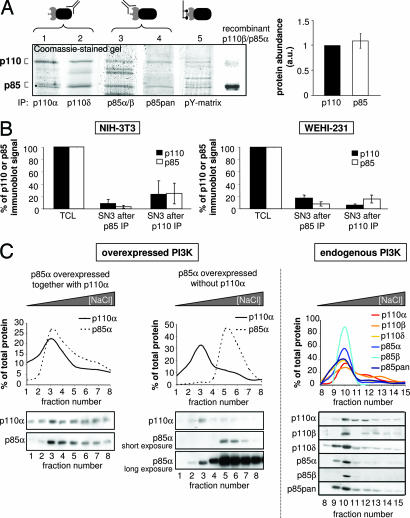
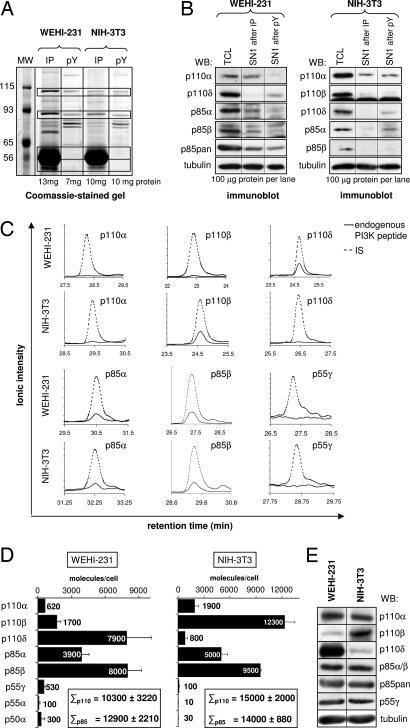
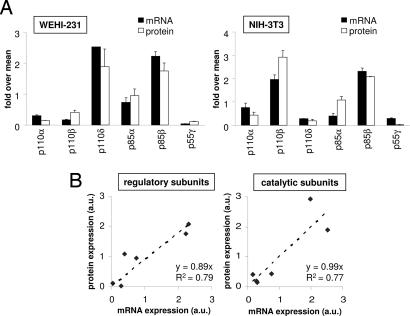
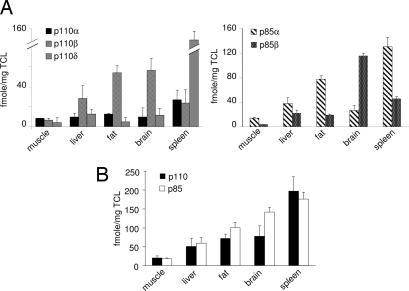
Class IA phosphoinositide 3-kinases (PI3Ks) signal downstream of tyrosine kinases and Ras and control a wide variety of biological responses. In mammals, these heterodimeric PI3Ks consist of a p110 catalytic subunit (p110alpha, p110beta, or p110delta) bound to any of five distinct regulatory subunits (p85alpha, p85beta, p55gamma, p55alpha, and p50alpha, collectively referred to as "p85s"). The relative expression levels of p85 and p110 have been invoked to explain key features of PI3K signaling. For example, free (i.e., non-p110-bound) p85alpha has been proposed to negatively regulate PI3K signaling by competition with p85/p110 for recruitment to phosphotyrosine docking sites. Using affinity and ion exchange chromatography and quantitative mass spectrometry, we demonstrate that the p85 and p110 subunits are present in equimolar amounts in mammalian cell lines and tissues. No evidence for free p85 or p110 subunits could be obtained. Cell lines contain 10,000-15,000 p85/p110 complexes per cell, with p110beta and p110delta being the most prevalent catalytic subunits in nonleukocytes and leukocytes, respectively. These results argue against a role of free p85 in PI3K signaling and provide insights into the nonredundant functions of the different class IA PI3K isoforms.
Conflict of interest statement
Conflict of interest statement: Bart Vanhaesebroeck is a consultant for PIramed (Slough, U.K.).
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
