

Mitotic spindle orientation distinguishes stem cell and terminal modes of neuron production in the early spinal cord
- PMID: 17470968
- PMCID: PMC7116174
- DOI: 10.1242/dev.002519
Mitotic spindle orientation distinguishes stem cell and terminal modes of neuron production in the early spinal cord
Abstract
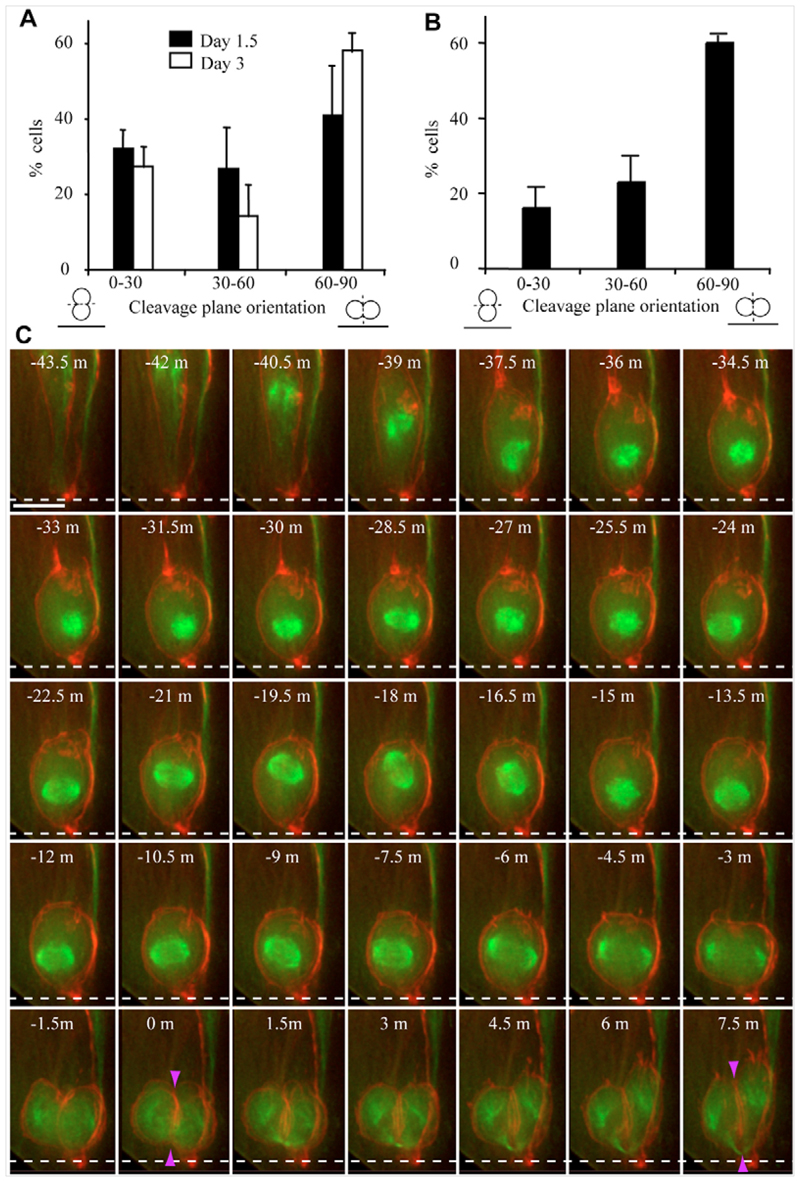
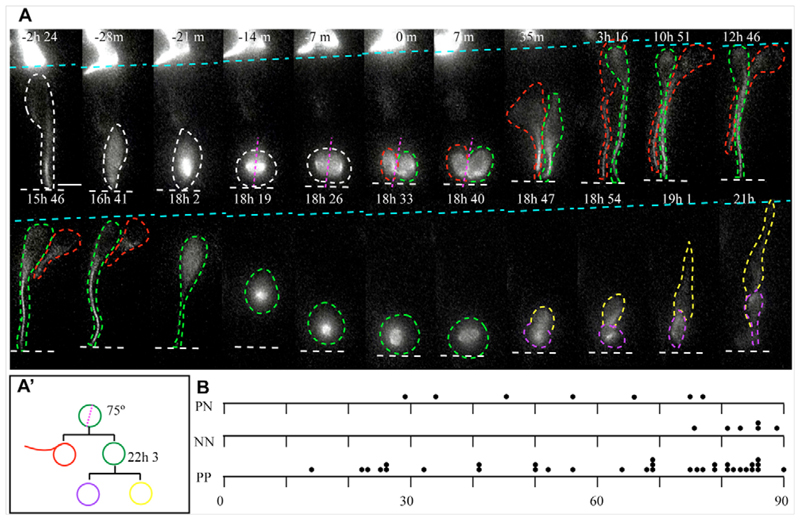
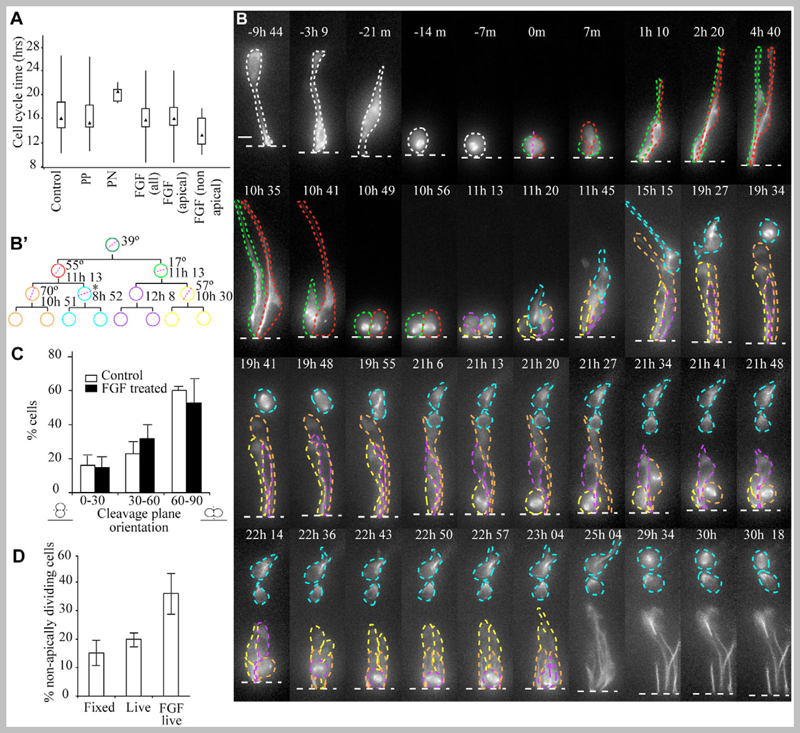
Despite great insight into the molecular mechanisms that specify neuronal cell type in the spinal cord, cell behaviour underlying neuron production in this tissue is largely unknown. In other neuroepithelia, divisions with a perpendicular cleavage plane at the apical surface generate symmetrical cell fates, whereas a parallel cleavage plane generates asymmetric daughters, a neuron and a progenitor in a stem cell mode, and has been linked to the acquisition of neuron-generating ability. Using a novel long-term imaging assay, we have monitored single cells in chick spinal cord as they transit mitosis and daughter cells become neurons or divide again. We reveal new morphologies accompanying neuron birth and show that neurons are generated concurrently by asymmetric and terminal symmetric divisions. Strikingly, divisions that generate two progenitors or a progenitor and a neuron both exhibit a wide range of cleavage plane orientations and only divisions that produce two neurons have an exclusively perpendicular orientation. Neuron-generating progenitors are also distinguished by lengthening cell cycle times, a finding supported by cell cycle acceleration on exposure to fibroblast growth factor (FGF), an inhibitor of neuronal differentiation. This study provides a novel, dynamic view of spinal cord neurogenesis and supports a model in which cleavage plane orientation/mitotic spindle position does not assign neuron-generating ability, but functions subsequent to this step to distinguish stem cell and terminal modes of neuron production.
Figures
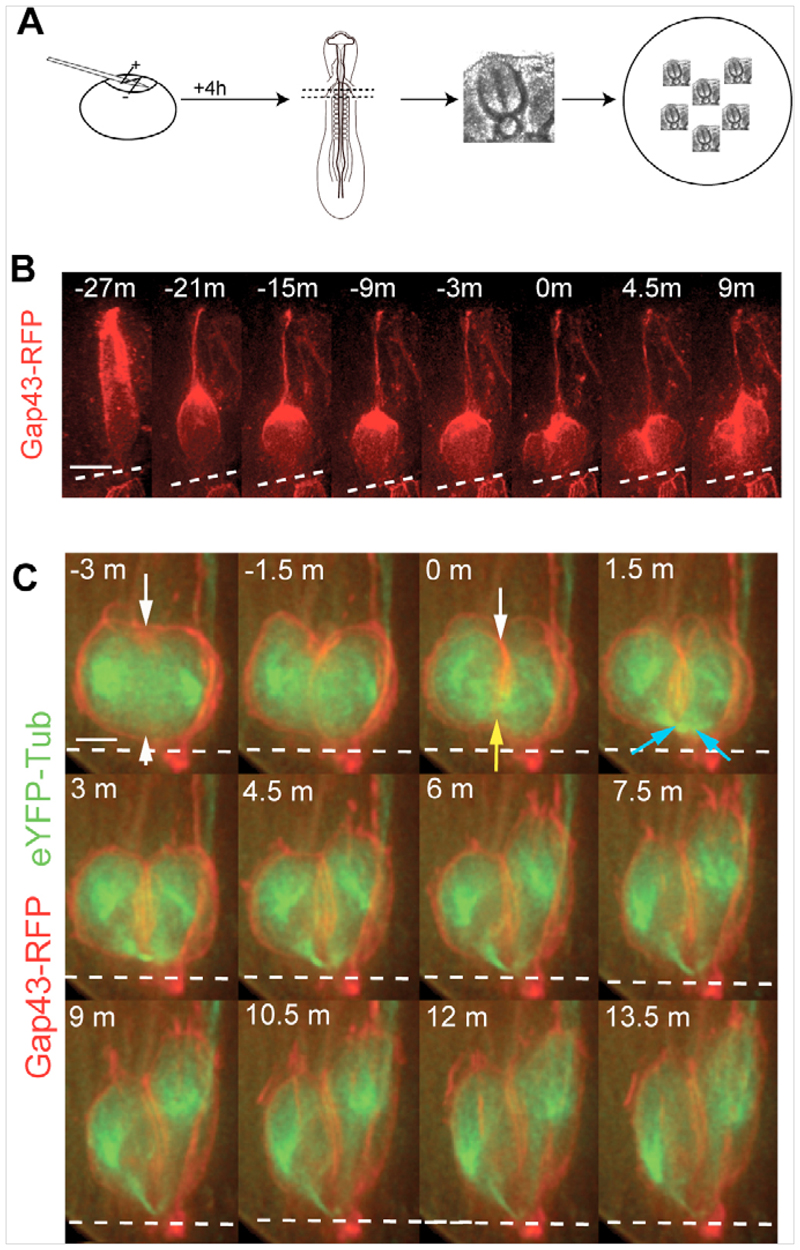
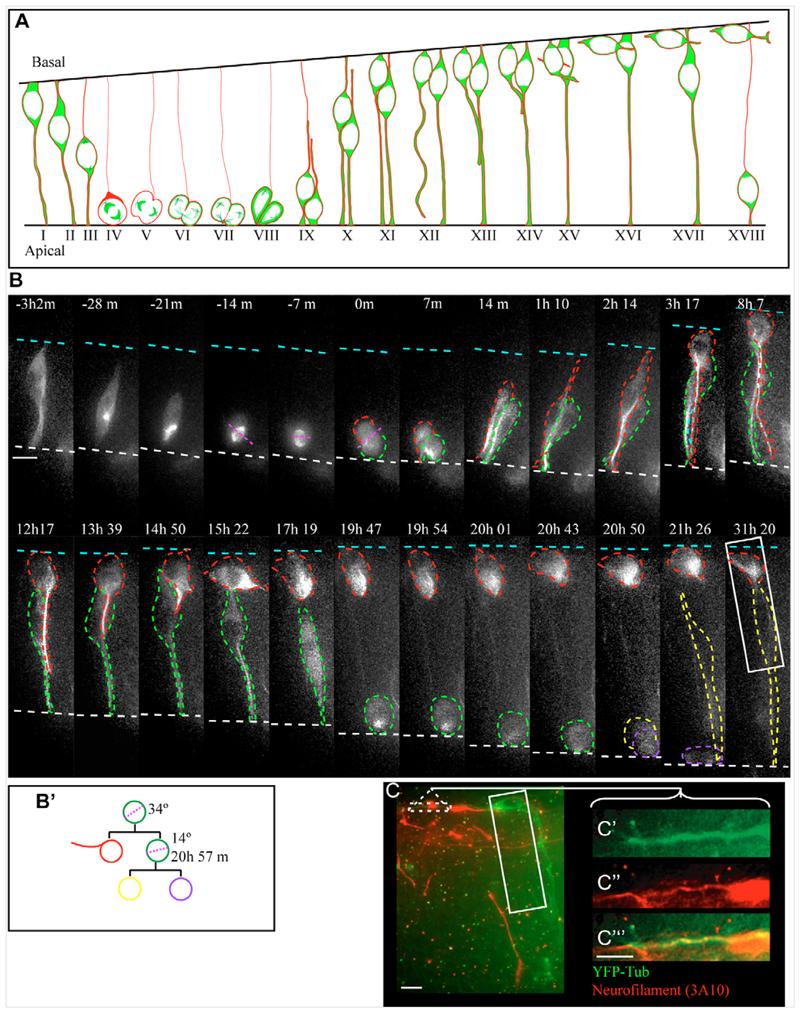
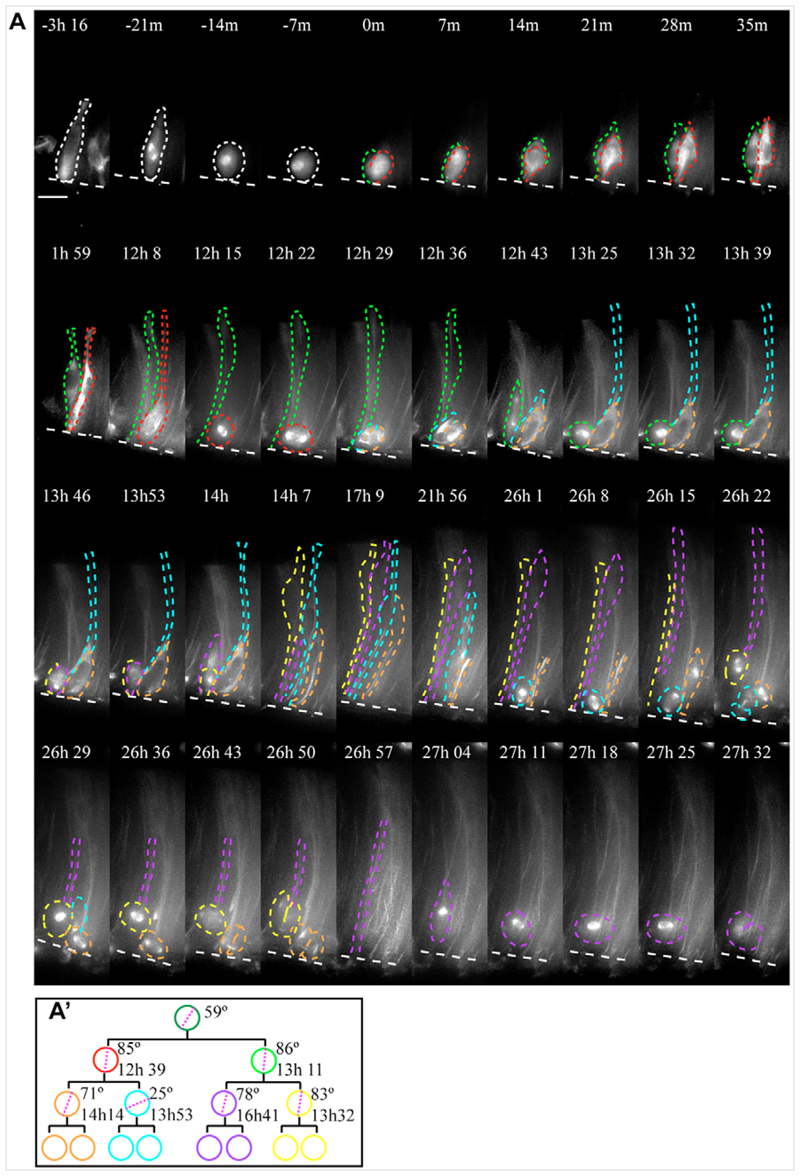
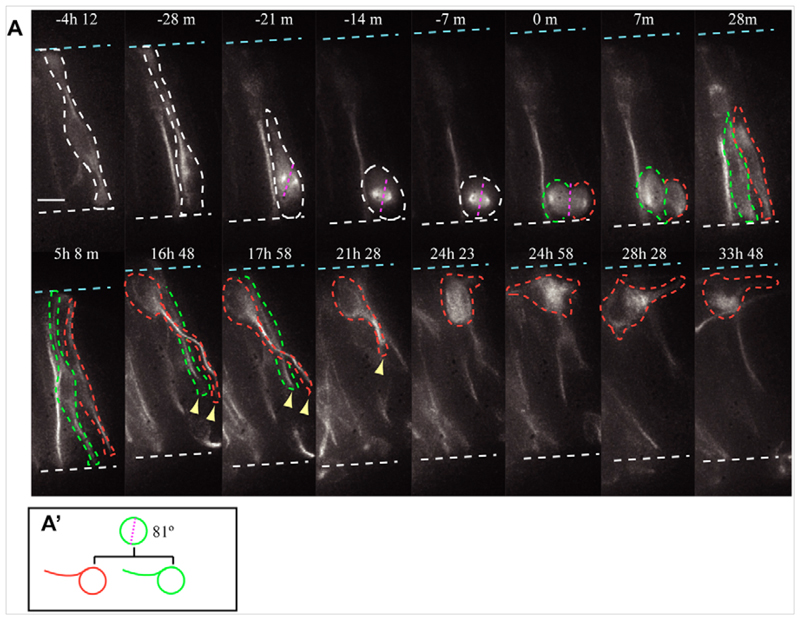
References
-
- Afonso C, Henrique D. PAR3 acts as a molecular organizer to define the apical domain of chick neuroepithelial cells. J Cell Sci. 2006;119:4293–4304. - PubMed
-
- Bertrand N, Castro DS, Guillemot F. Proneural genes and the specification of neural cell types. Nat Rev Neurosci. 2002;3:517–530. - PubMed
-
- Betschinger J, Knoblich JA. Dare to be different: asymmetric cell division in Drosophila, C. elegans and vertebrates. Curr Biol. 2004;14:R674–R685. - PubMed
-
- Cai L, Hayes NL, Takahashi T, Caviness VS, Jr, Nowakowski RS. Size distribution of retrovirally marked lineages matches prediction from population measurements of cell cycle behavior. J Neurosci Res. 2002;69:731–744. - PubMed
-
- Calegari F, Huttner WB. An inhibition of cyclin-dependent kinases that lengthens, but does not arrest, neuroepithelial cell cycle induces premature neurogenesis. J Cell Sci. 2003;116:4947–4955. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
