

Review
doi: 10.1186/gb-2007-8-4-210.
Searching genomes for ribozymes and riboswitches
Affiliations
- PMID: 17472738
- PMCID: PMC1895996
- DOI: 10.1186/gb-2007-8-4-210
Item in Clipboard
Review
Searching genomes for ribozymes and riboswitches
Genome Biol.
2007.
Abstract
New regulatory RNAs with complex structures have recently been discovered, among them the first catalytic riboswitch, a gene-regulatory RNA sequence with catalytic activity. Here we discuss some of the experimental approaches and theoretical difficulties attached to the identification of new ribozymes in genomes.
Figures
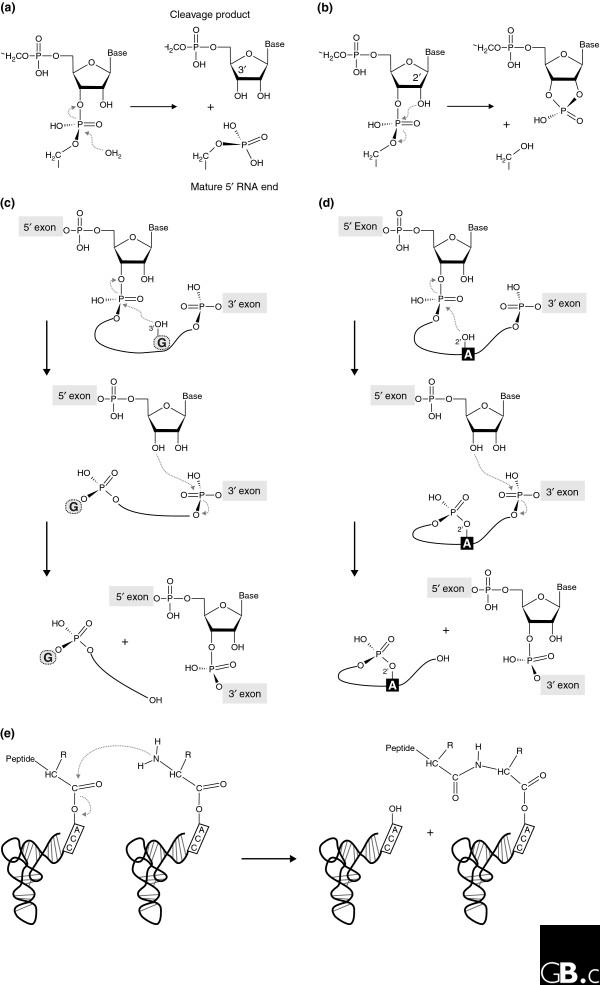
Biochemical reactions naturally catalyzed by RNA. (a) Precursor tRNA hydrolysis by bacterial RNase P yields a phosphate-containing 5' end of the mature tRNA and a 3'-hydroxyl group at the 5' cleavage product. (b-d) Transesterification reactions catalyzed by (b) the small nucleolytic ribozymes, (c) group I introns, and (d) group II introns, in which different chemical groups serve as the attacking nucleophile. In the small nucleolytic ribozymes (b), a defined 2'-hydroxyl attacks the neighboring 3',5'-phosphodiester bond, resulting in a 2',3'-cyclic phosphate and a 5'-hydroxyl in the respective cleavage products. In the first step of group I intron splicing (c), the 3'-hydroxyl of the exogenous guanosine (G) cofactor attacks the 5'-exon-intron junction and sets the 5' exon free, which leads to the covalent attachment of the cofactor to the 5' end of the intron. In a second transesterification reaction, the 5' exon forms a conventional 3',5' bond with the 3' exon, releasing the linear intron with the additional guanosine [1]. In group II introns (d), the conserved branch-point adenosine (A) serves as the nucleophile, leading to the formation of a lariat intron. (e) Peptide-bond formation catalyzed by the ribosome.
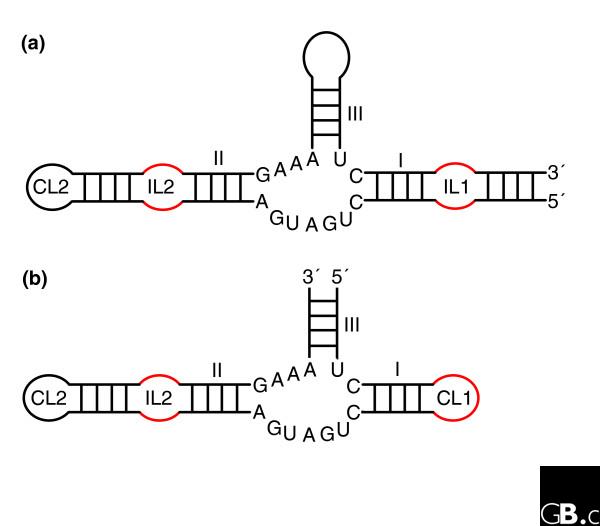
The hammerhead ribozymes are based on a three-way junction and there are two main types. (a) Type I has the ends of the single-stranded RNA on stem I; (b) type III has the ends of the single-stranded RNA on stem III. For unknown reasons, potential type II ribozymes (ends of the single-stranded RNA on stem II) have never been observed. The three-dimensional architecture is maintained by coaxial stacking of stems II and III, which, through constraints in the conserved three-way junction residues [92], orients stem I so that loop-loop interactions between stems I and II form (Figure 3) [40,42]. The internal loop of stem II (IL2) is often replaced by a capping loop (CL2); similarly, CL1 in type III can be replaced by an internal loop (IL1) followed by another hairpin. Although only one structure has been fully characterized, sequence alignments show that the loop-loop interactions (mainly constituting non-Watson-Crick pairs) are very diverse.
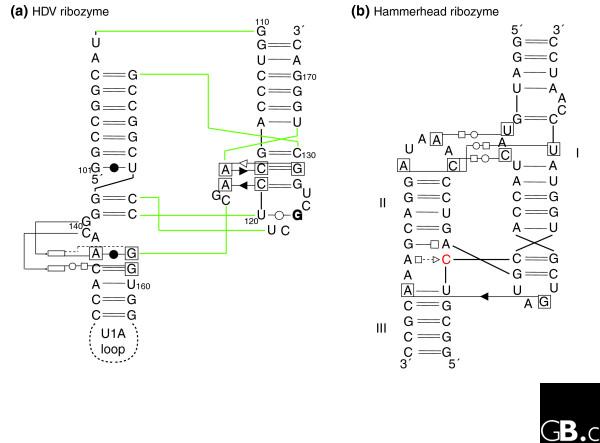
Schematic diagrams of the interaction networks maintaining the three-dimensional architecture of two different ribozymes. (a) The HDV ribozyme [7,93]; (b) the active hammerhead ribozyme [42]. The HDV ribozyme has a convoluted pseudoknotted topology: the color lines indicate the path of the sugar-phosphate backbone. The nomenclature is as follows [75]. Each nucleotide has three edges with hydrogen bonding possibilities: the Watson-Crick edge (denoted by a circle), the Hoogsteen edge (denoted by a square) and the sugar edge (denoted by a triangle). A pairwise base-base interaction can be formed either with the attached sugar moieties on the same side of the line of approach (cis-configuration, the symbols are closed) or with the sugars on either sides of the line of approach (the trans-configuration, the symbols are open). To avoid ambiguities, when annotating tertiary contacts, the nucleotides that are involved have been boxed. When the base of a nucleotide is in the syn-conformation with respect to the sugar it is marked in bold. The rectangles indicate the position actually occupied in space by a nucleotide. In (b), the cleavage occurs 3' of the red C.
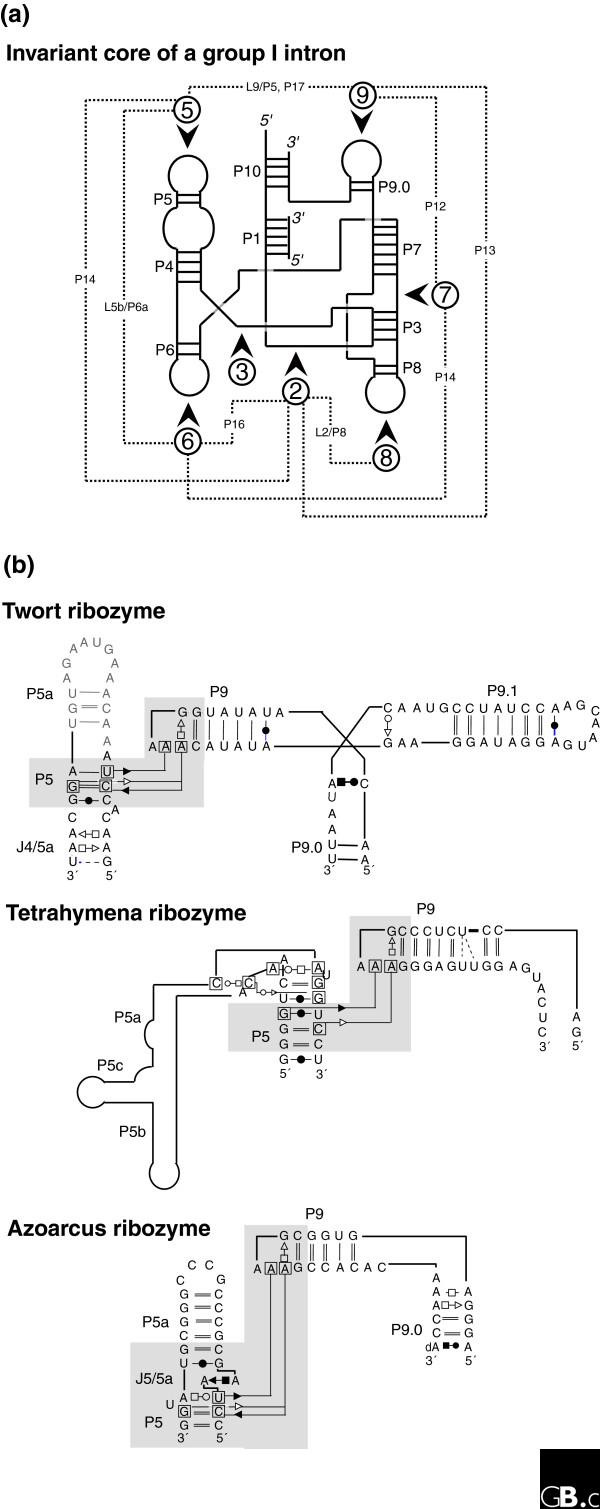
Different local topologies can give rise to similar tertiary contacts in group I introns. (a) The invariant core of a group I intron [36,94] is illustrated in schematic form with the paired segments indicated by P and the loop regions by L. The dashed lines indicate the contacts between the peripheral elements, which are indicated by the numbers in circles. (b) Three different group I introns illustrate distinct ways of achieving a similar tertiary contact (involving non-Watson-Crick A-minor base-base interactions between a GAAA tetraloop and two stacked pairs) connecting distant regions. In each case region 9 folds towards region 5 (as indicated by the shaded region) but, in the Twort ribozyme [95] this is via a three-way junction, in the Tetrahymena ribozyme [96], it is via a large bend (this is not the natural junction, however), and in the Azoarcus ribozyme [97], it is via a kink-turn. Each motif has a different sequence and set of structural constraints [77,92].
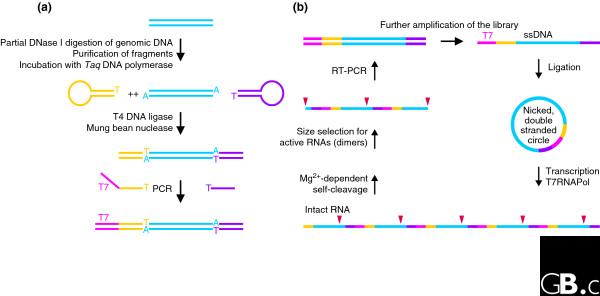
Identification of catalytic RNA from a genomic library. (a) Preparation of the genomic library. Genomic DNA is first partially digested and fragments of approximately 150 bp (blue) are gel-purified and incubated with Taq polymerase to give them 3' A overhangs. In the next step, ligation of covalently closed oligonucleotides (yellow and purple) to the library prevents the unwanted combination of DNA fragments. After removal of DNA hairpins, a T7 promoter (magenta) is then added by PCR, yielding an amplified linear library. (b) The in vitro selection scheme. The library is further amplified by PCR using a 5'-phosphorylated reverse primer and a biotinylated forward primer that allows the isolation of the phosphorylated strand using streptavidin beads. Single strands are individually circularized by ligation with a splint oligonucleotide and the second strand is added by incubation with Taq polymerase and deoxynucleoside triphosphates. The resulting nicked double-stranded library is suitable for rolling-circle transcription by T7 polymerase [98], yielding multimeric RNA species potentially encoding sites of self-cleavage (red triangles). The RNA is then incubated for self-cleavage, and active molecules (dimers) are size-selected. The scheme is completed by preparation of the next-generation DNA library using reverse transcription-PCR (RT-PCR). Modified from [29].
References
-
- Buzayan JM, Gerlach WL, Bruening G. Non-enzymatic cleavage and ligation of RNAs complementary to a plant virus satellite RNA. Nature. 1986;323:349–353. doi: 10.1038/323349a0. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
