

Review
doi: 10.1038/nrm2178.
RNA-binding proteins: modular design for efficient function
Affiliations
- PMID: 17473849
- PMCID: PMC5507177
- DOI: 10.1038/nrm2178
Item in Clipboard
Review
RNA-binding proteins: modular design for efficient function
Nat Rev Mol Cell Biol.
2007 Jun.
Abstract
Many RNA-binding proteins have modular structures and are composed of multiple repeats of just a few basic domains that are arranged in various ways to satisfy their diverse functional requirements. Recent studies have investigated how different modules cooperate in regulating the RNA-binding specificity and the biological activity of these proteins. They have also investigated how multiple modules cooperate with enzymatic domains to regulate the catalytic activity of enzymes that act on RNA. These studies have shown how, for many RNA-binding proteins, multiple modules define the fundamental structural unit that is responsible for biological function.
Figures
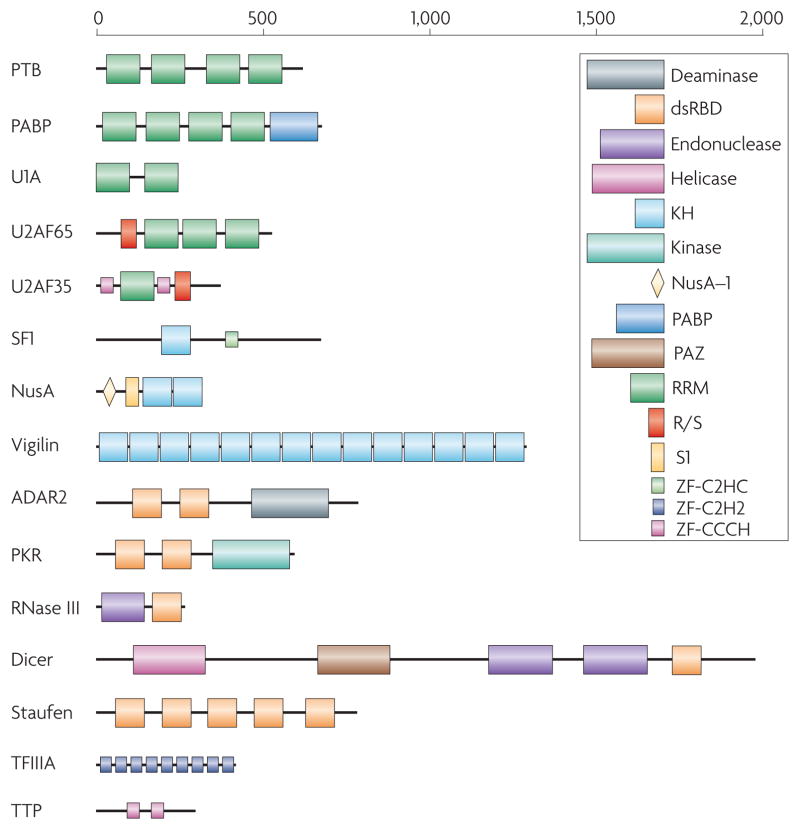
Representative examples from some of the most common RNA-binding protein families, as illustrated here, demonstrate the variability in the number of copies (as many as 14 in vigilin) and arrangements that exist in RNA-binding proteins. This variability has direct functional implications. For example, Dicer and RNase III both contain an endonuclease catalytic domain followed by an RNA-binding dsRBD; thus, both proteins recognize double-stranded RNA, but Dicer has evolved to interact specifically with RNA species produced through the RNA interference pathway through additional domains that recognize the unique structural features of these RNAs. Different domains are schematically represented in colored boxes, including the RRM (RNA Recognition Motif; green), by far the most common RNA-binding protein module; the KH (K-Homology) domain, blue, capable of binding both single stranded RNA and DNA; the dsRBD (double-stranded RNA-binding domain), a sequence-independent dsRNA binding module (red); RNA-binding zinc finger domains (light blue or pink). Enzymatic domains and less common functional modules are indicated in a variety of colors.
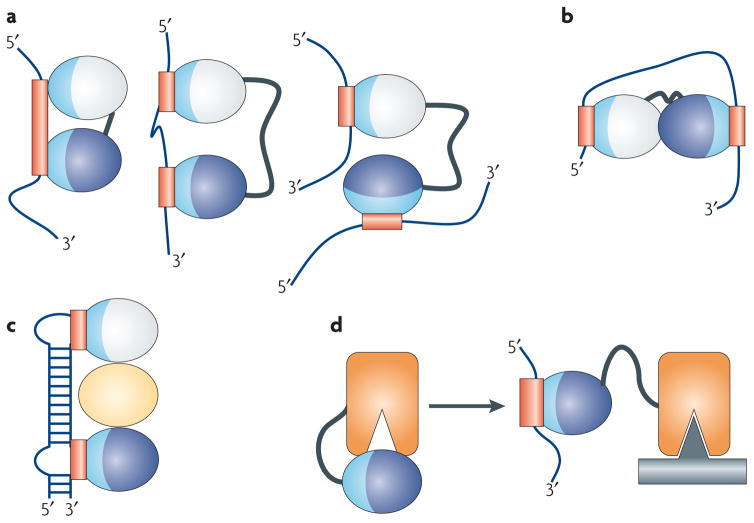
RNA-binding domains function in a variety of ways. (a) They recognize RNA sequences with a specificity and affinity that would not be possible through a single domain or if multiple domains did not cooperate. Multiple domains combine to recognize a longer RNA sequence (left), sequences separated by many nucleotides (centre), or RNAs belonging to different molecules altogether (right). (b) RNA-binding domains can organize mRNAs topologically by interacting simultaneously with multiple RNA sequences or (c) they can function as spacers to properly position other modules for recognition. (d) They can combine with enzymatic domains to define the substrate specificity for catalysis or regulate enzymatic activity. In this figure, the RNA-binding modules are represented as ellipses with their RNA-binding surfaces colored in light blue, with the corresponding binding sites within the RNA colored in red; individual domains are colored differently.
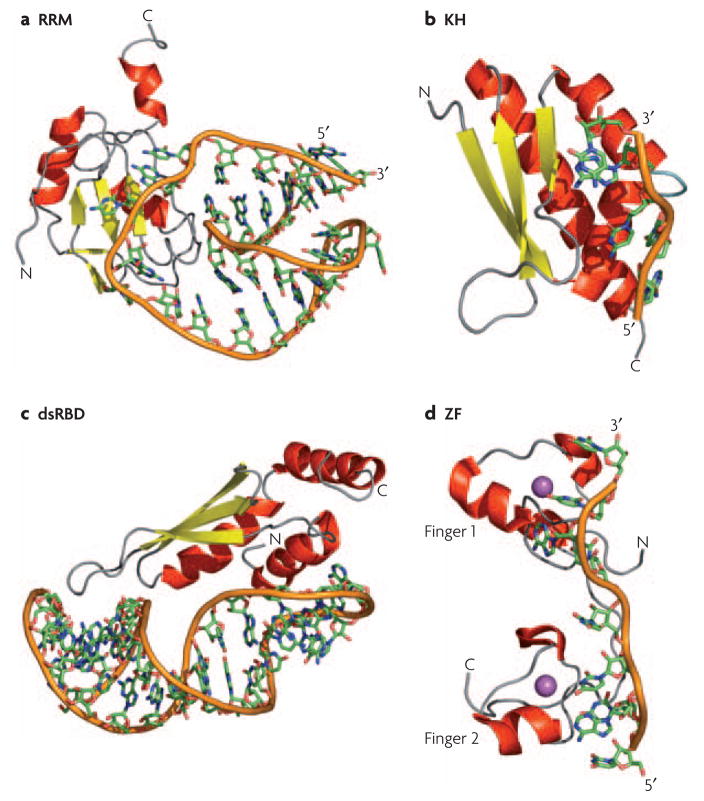
a) Structure of the N-terminal RNA-recognition motif (RRM) of human U1A bound to RNA; in this structure, and in many other RRM-RNA complexes, single stranded bases are specifically recognized through the protein β-sheet and two loops connecting the secondary structure elements. b) The hnRNP K homology 3 (KH3) domain of Nova-2 bound to 5′-AUCAC-3′; these domains bind to both single-stranded DNA and RNA through a conserved GXXG sequence located in an exposed loop (light blue). c) The Rnt1 double-stranded RNA-binding domain (dsRBD) bound to a RNA helix capped by an AGNN tetraloop; a conserved protein loop (left-most part of the structure) interacts with 2′-OH groups in the RNA minor groove while highly conserved Lys and Arg residues at the end of the longer helix recognize the position of phosphate atoms characteristic of an A-form helix. d) The two zinc fingers of Tis11d bound to an AU-rich RNA element; the identity of the single stranded RNA is recognized by the protein backbone through hydrogen bonds with the Watson-Crick face of each base. In all panels, the RNA backbone is represented with an orange ribbon, α-helices are in red and β-sheets in yellow; the Zn atom in the Tis11d structure is in magenta.
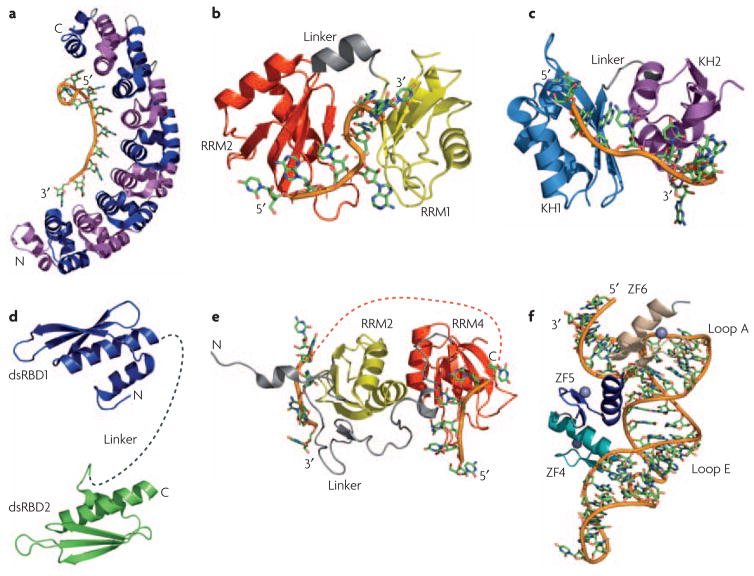
a) The structure of human Pumilio provides an example of how multiple repeats (eight in this case) that individually recognize a few nucleotides (one in this case) combine to specifically recognize a much longer RNA sequence. Repeats are alternatively colored in magenta and blue; the RNA is colored similarly in all other structures with the backbone shown as an orange tube. b) In the structures of the two RNA recognition motifs (RRMs) from Hrp1 and c) the two K-homology (KH) domains of NusA, a short linker (in gray) allows the two domains to position themselves with respect to one another upon binding RNA; for Hrp1, the first RRM is in yellow and the second in red; for NusA, the first KH domain is cyan and the second is purple. d) Flexibility within the linker between two double-stranded RNA-binding domains (dsRBDs) allow recognition of separated binding sites. The two dsRBDs of ADAR2 are connected by a flexible linker (dashed gray line) that may allow the protein to interact with a variety of targets of different structure. e) The RNA-recognition motifs RRM3 (yellow) and RRM4 (red) of PTB form interdomain interactions involving the face of the protein opposite to the β-sheet involved in RNA recognition. This interaction positions the two domains in such away that interacting RNA sequences are looped away from each other, as indicated by the orange dotted line connecting the two RNAs. f) The structure of the TFIIIA-RNA complex illustrates how zinc finger ZF5 (blue) functions as a spacer that properly positions zinc fingers 4 (teal) and 6 (tan) for recognition of loops E and A, respectively, within 5S rRNA.
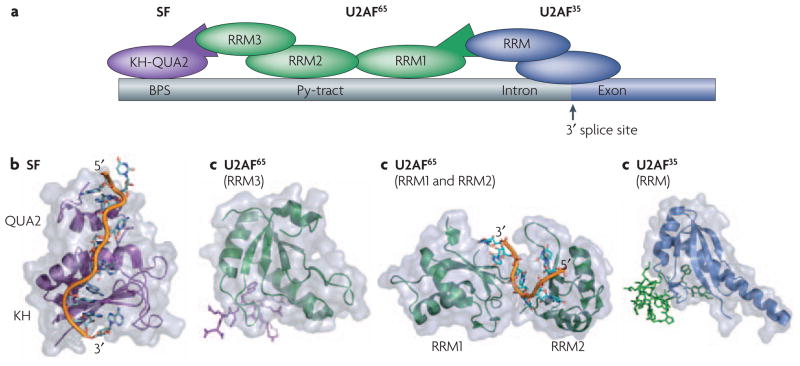
Proper definition of the splicing site requires a number of cooperative binding events that are mediated by both protein-protein and protein-RNA contacts between various RNA binding modules. a) Schematic of the interactions between various proteins and RNA at the splicing site. Some of the key domains involved in these interactions whose structures are shown below are labeled. Within the RNA the branch-point sequence (BPS), pyrimidine tract (Py-tract), and the 3′ splice site (3′ ss) are labeled with the intron colored gray and the exon colored dark blue. b) SF1 recognizes the BPS through its KH-QUA2 domains. The additional QUA2 domain creates an extended KH domain that can recognize the full BPS sequence RNA. c) This interaction is strengthened by protein-protein interactions between it’s N-terminus and the non-canonical RRM3 of U2AF6535, d) which is bound to the pyrmidine tract through it’s first two canonical RRMs. Finally, the U2AF interaction is also aided by protein-protein interactions between it’s N-terminus and the non-canonical RRM of U2AF bound at the 3′ splice site. All of the protein and peptide structures are colored as shown in the schematic (a).
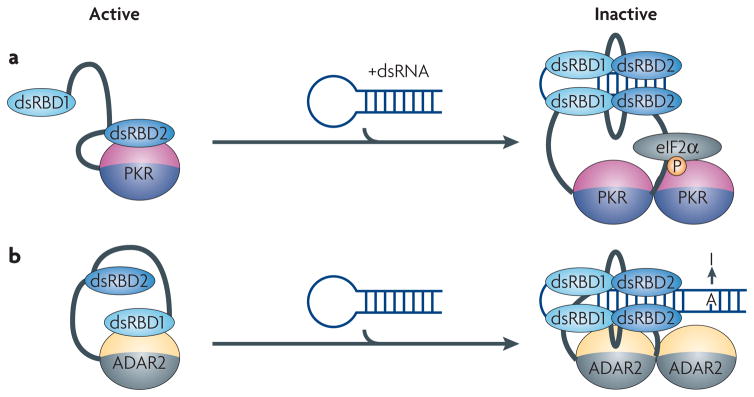
In both PKR and ADAR proteins, inter-domain interactions between the RNA-binding module and the catalytic domain maintain the proteins in an inactive state. a) The kinase domain of PKR is inhibited by an interaction with the double-stranded RNA-binding domain (dsRBD2). Binding to dsRNA releases the kinase from its inactive state allowing it to inhibit translation by phosphorylating eIF2α. b) The activity of ADAR2 is controlled by a mechanism similar to PKR, but in this case dsRBD1 is involved in the inactivation of the catalytic domain. When double-stranded RNA (dsRNA) binds to both dsRBDs, the protein dimerizes and the catalytic domain becomes exposed to convert adenosine to inosine.
References
-
- Dreyfuss G, Kim VN, Kataoka N. Messenger-RNA-binding proteins and the messages they carry. Nat Rev Mol Cell Biol. 2002;3:195–205. - PubMed
-
- Burd CG, Dreyfuss G. Conserved structures and diversity of functions of RNA-binding proteins. Science. 1994;265:615–21. - PubMed
-
- Chang KY, Ramos A. The double-stranded RNA-binding motif, a versatile macromolecular docking platform. Febs J. 2005;272:2109–17. - PubMed
-
- Hall TM. Multiple modes of RNA recognition by zinc finger proteins. Curr Opin Struct Biol. 2005;15:367–73. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
