

Drosophila as a genetic and cellular model for studies on axonal growth
- PMID: 17475018
- PMCID: PMC1876224
- DOI: 10.1186/1749-8104-2-9
Drosophila as a genetic and cellular model for studies on axonal growth
Abstract
One of the most fascinating processes during nervous system development is the establishment of stereotypic neuronal networks. An essential step in this process is the outgrowth and precise navigation (pathfinding) of axons and dendrites towards their synaptic partner cells. This phenomenon was first described more than a century ago and, over the past decades, increasing insights have been gained into the cellular and molecular mechanisms regulating neuronal growth and navigation. Progress in this area has been greatly assisted by the use of simple and genetically tractable invertebrate model systems, such as the fruit fly Drosophila melanogaster. This review is dedicated to Drosophila as a genetic and cellular model to study axonal growth and demonstrates how it can and has been used for this research. We describe the various cellular systems of Drosophila used for such studies, insights into axonal growth cones and their cytoskeletal dynamics, and summarise identified molecular signalling pathways required for growth cone navigation, with particular focus on pathfinding decisions in the ventral nerve cord of Drosophila embryos. These Drosophila-specific aspects are viewed in the general context of our current knowledge about neuronal growth.
Figures
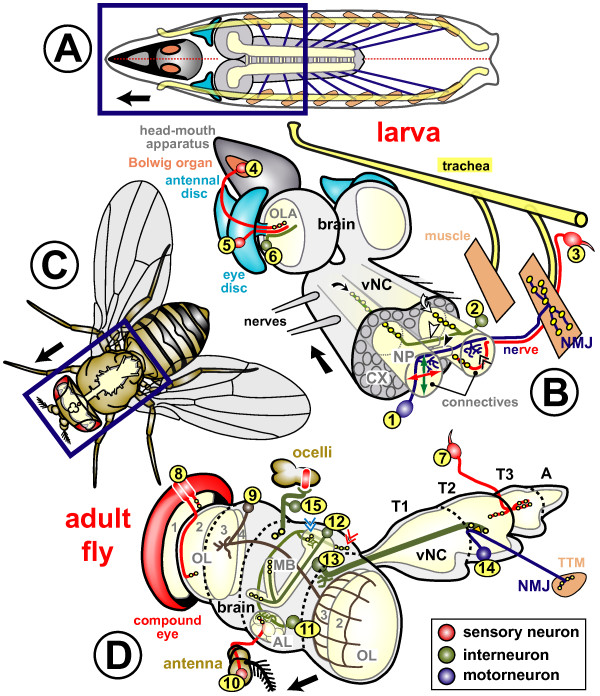
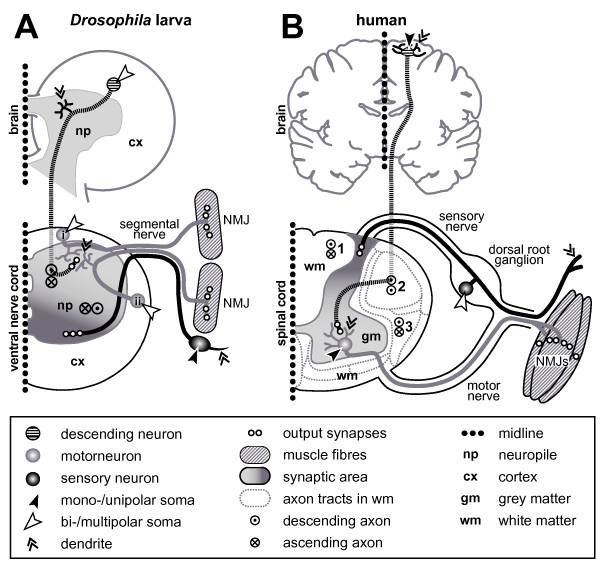
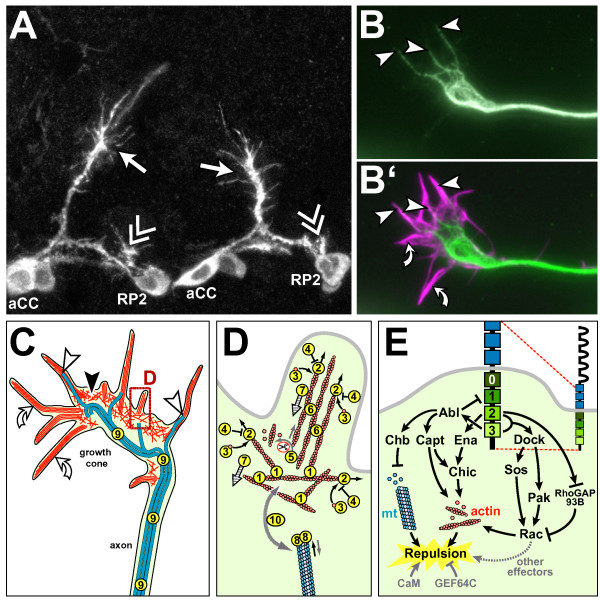

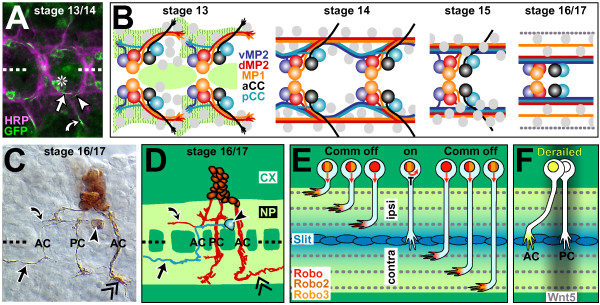
References
-
- Ramón y Cajal S. Histology of the Nervous System of Man and Vertebrates (English translation by Swanson N, Swanson LW from 1995) New York Oxford: Oxford University Press; 1909. p. 1911.
-
- Tessier-Lavigne M, Goodman CS. The molecular biology of axon guidance. Science. 1996;274:1123–1133. - PubMed
-
- Araujo SJ, Tear G. Axon guidance mechanisms and molecules: lessons from invertebrates. Nat Rev Neurosci. 2003;4:910–922. - PubMed
-
- Bate CM. Pioneer neurones in an insect embryo. Nature. 1976;260:54–56. - PubMed
-
- Harrison RG. The outgrowth of the nerve fiber as a mode of protoplasmic movement. J Exp Zool. 1910;9:787–846. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
