

Overexpressing centriole-replication proteins in vivo induces centriole overduplication and de novo formation
- PMID: 17475495
- PMCID: PMC1885955
- DOI: 10.1016/j.cub.2007.04.036
Overexpressing centriole-replication proteins in vivo induces centriole overduplication and de novo formation
Abstract
Background: Centrosomes have important roles in many aspects of cell organization, and aberrations in their number and function are associated with various diseases, including cancer. Centrosomes consist of a pair of centrioles surrounded by a pericentriolar matrix (PCM), and their replication is tightly regulated. Here, we investigate the effects of overexpressing the three proteins known to be required for centriole replication in Drosophila-DSas-6, DSas-4, and Sak.
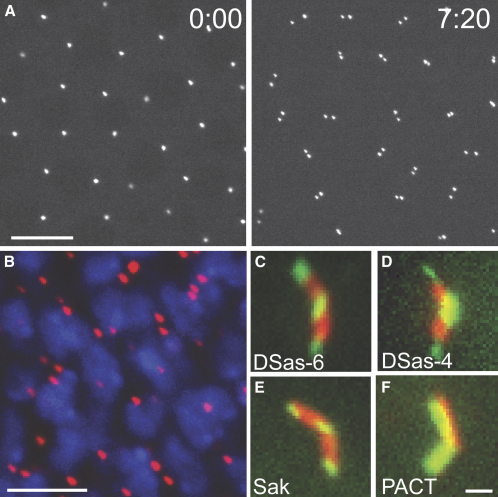
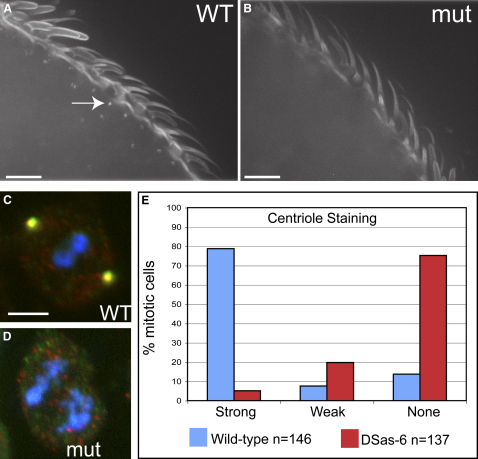
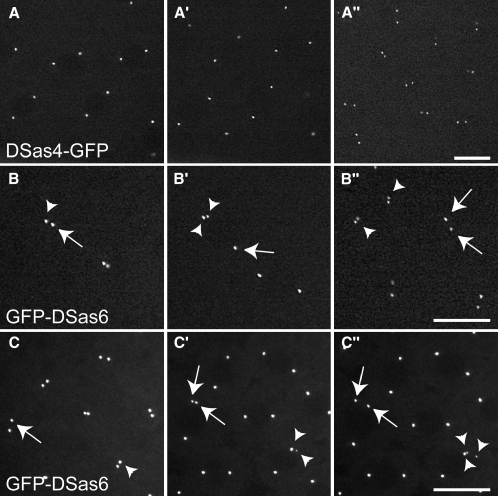
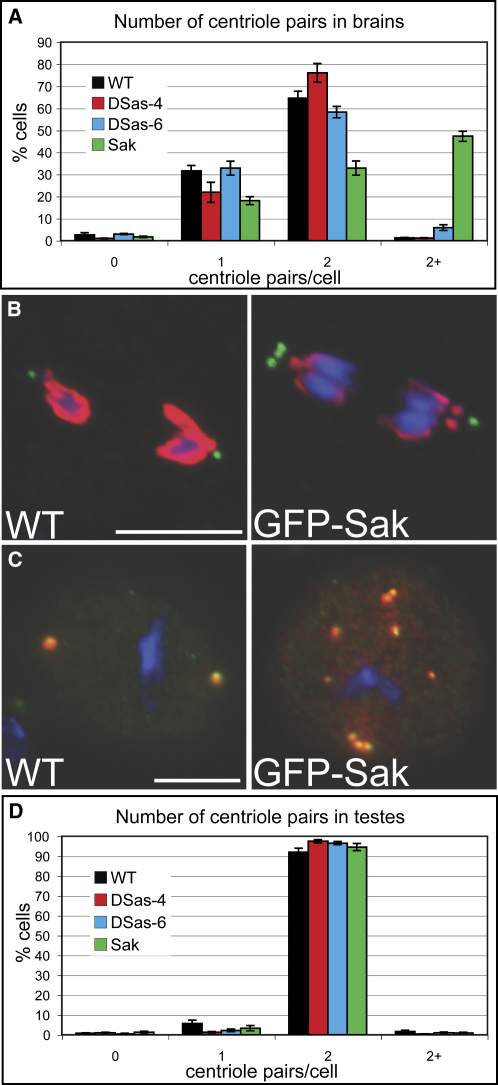
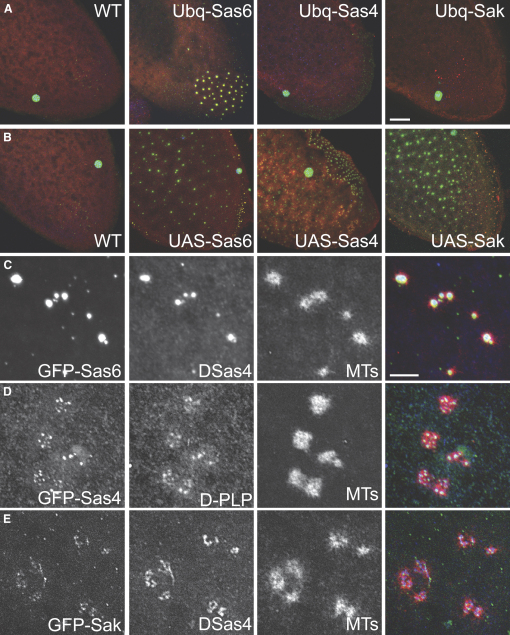
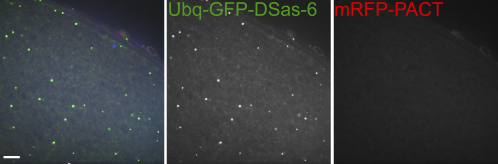
Results: By directly observing centriole replication in living Drosophila embryos, we show that the overexpression of GFP-DSas-6 can drive extra rounds of centriole replication within a single cell cycle. Extra centriole-like structures also accumulate in brain cells that overexpress either GFP-DSas-6 or GFP-Sak, but not DSas-4-GFP. No extra centrioles accumulate in spermatocytes that overexpress any of these three proteins. Most remarkably, the overexpression of any one of these three proteins results in the rapid de novo formation of many hundreds of centriole-like structures in unfertilized eggs, which normally do not contain centrioles.
Conclusions: Our data suggest that the levels of centriolar DSas-6 determine the number of daughter centrioles formed during centriole replication. Overexpression of either DSas-6 or Sak can induce the formation of extra centrioles in some tissues but not others, suggesting that centriole replication is regulated differently in different tissues. The finding that the overexpression of DSas-4, DSas-6, or Sak can rapidly induce the de novo formation of centriole-like structures in Drosophila eggs suggests that this process results from the stabilization of centriole-precursors that are normally present in the egg.
Figures
References
-
- Kellogg D.R., Moritz M., Alberts B.M. The centrosome and cellular organization. Annu. Rev. Biochem. 1994;63:639–674. - PubMed
-
- Nigg E.A. Centrosome aberrations: cause or consequence of cancer progression? Nat. Rev. Cancer. 2002;2:815–825. - PubMed
-
- Raff J.W. Centrosomes and cancer: lessons from a TACC. Trends Cell Biol. 2002;12:222–225. - PubMed
-
- Basto R., Lau J., Vinogradova T., Gardiol A., Woods C.G., Khodjakov A., Raff J.W. Flies without centrioles. Cell. 2006;125:1375–1386. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
