

A new time-scale for ray-finned fish evolution
- PMID: 17476768
- PMCID: PMC1766393
- DOI: 10.1098/rspb.2006.3749
A new time-scale for ray-finned fish evolution
Abstract
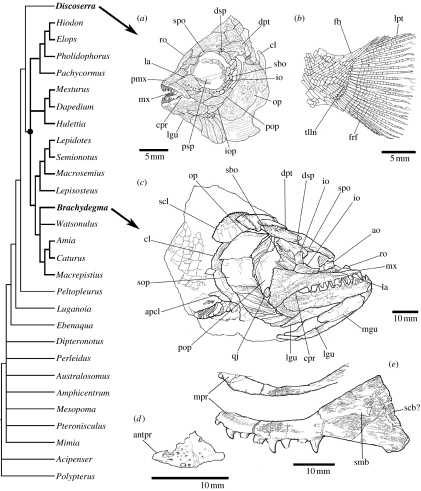
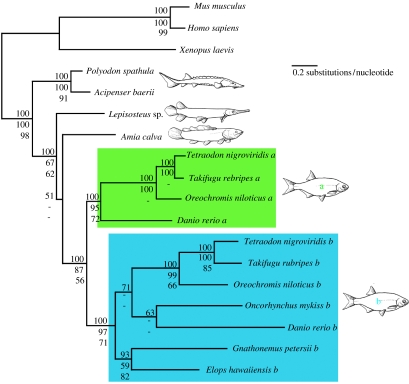
The Actinopterygii (ray-finned fishes) is the largest and most diverse vertebrate group, but little is agreed about the timing of its early evolution. Estimates using mitochondrial genomic data suggest that the major actinopterygian clades are much older than divergence dates implied by fossils. Here, the timing of the evolutionary origins of these clades is reinvestigated using morphological, and nuclear and mitochondrial genetic data. Results indicate that existing fossil-based estimates of the age of the crown-group Neopterygii, including the teleosts, Lepisosteus (gar) and Amia (bowfin), are at least 40 Myr too young. We present new palaeontological evidence that the neopterygian crown radiation is a Palaeozoic event, and demonstrate that conflicts between molecular and morphological data for the age of the Neopterygii result, in part, from missing fossil data. Although our molecular data also provide an older age estimate for the teleost crown, this range extension remains unsupported by the fossil evidence. Nuclear data from all relevant clades are used to demonstrate that the actinopterygian whole-genome duplication event is teleost-specific. While the date estimate of this event overlaps the probable range of the teleost stem group, a correlation between the genome duplication and the large-scale pattern of actinopterygian phylogeny remains elusive.
Figures


Similar articles
-
Major issues in the origins of ray-finned fish (Actinopterygii) biodiversity.Biol Rev Camb Philos Soc. 2014 Nov;89(4):950-71. doi: 10.1111/brv.12086. Epub 2014 Feb 24. Biol Rev Camb Philos Soc. 2014. PMID: 24612207
-
Early members of 'living fossil' lineage imply later origin of modern ray-finned fishes.Nature. 2017 Sep 14;549(7671):265-268. doi: 10.1038/nature23654. Epub 2017 Aug 30. Nature. 2017. PMID: 28854173
-
Little evidence for enhanced phenotypic evolution in early teleosts relative to their living fossil sister group.Proc Natl Acad Sci U S A. 2016 Oct 11;113(41):11531-11536. doi: 10.1073/pnas.1607237113. Epub 2016 Sep 26. Proc Natl Acad Sci U S A. 2016. PMID: 27671652 Free PMC article.
-
Vicariance and dispersal in southern hemisphere freshwater fish clades: a palaeontological perspective.Biol Rev Camb Philos Soc. 2019 Apr;94(2):662-699. doi: 10.1111/brv.12473. Epub 2018 Oct 19. Biol Rev Camb Philos Soc. 2019. PMID: 30338909 Review.
-
'Fish' (Actinopterygii and Elasmobranchii) diversification patterns through deep time.Biol Rev Camb Philos Soc. 2016 Nov;91(4):950-981. doi: 10.1111/brv.12203. Epub 2015 Jun 23. Biol Rev Camb Philos Soc. 2016. PMID: 26105527 Review.
Cited by
-
Comparable ages for the independent origins of electrogenesis in African and South American weakly electric fishes.PLoS One. 2012;7(5):e36287. doi: 10.1371/journal.pone.0036287. Epub 2012 May 14. PLoS One. 2012. PMID: 22606250 Free PMC article.
-
Did genome duplication drive the origin of teleosts? A comparative study of diversification in ray-finned fishes.BMC Evol Biol. 2009 Aug 8;9:194. doi: 10.1186/1471-2148-9-194. BMC Evol Biol. 2009. PMID: 19664233 Free PMC article.
-
Old gene duplication facilitates origin and diversification of an innovative communication system--twice.Proc Natl Acad Sci U S A. 2010 Dec 21;107(51):22172-7. doi: 10.1073/pnas.1011803107. Epub 2010 Dec 2. Proc Natl Acad Sci U S A. 2010. PMID: 21127261 Free PMC article.
-
The house spider genome reveals an ancient whole-genome duplication during arachnid evolution.BMC Biol. 2017 Jul 31;15(1):62. doi: 10.1186/s12915-017-0399-x. BMC Biol. 2017. PMID: 28756775 Free PMC article.
-
Distinct phenotypes in zebrafish models of human startle disease.Neurobiol Dis. 2013 Dec;60:139-51. doi: 10.1016/j.nbd.2013.09.002. Epub 2013 Sep 9. Neurobiol Dis. 2013. PMID: 24029548 Free PMC article.
References
-
- Amores A, et al. Zebrafish hox clusters and vertebrate genome evolution. Science. 1998;282:1711–1714. - PubMed
-
- Arratia G. Basal teleosts and teleostean phylogeny. Palaeo Ichthyologica. 1997;7:1–168.
-
- Arratia G. Remarkable teleostean fishes from the Late Jurassic of southern Germany and their phylogenetic relationships. Mitt. Mus. Nat.kd., Berl., Geowiss. Reihe. 2000;3:137–179.
-
- Cavin L, Suteethorn V. A new semionotiform (Actinopterygii, Neopterygii) from Upper Jurassic—Lower Cretaceous deposits of north-east Thailand, with comments on the relationships of semionotiforms. Palaeontology. 2006;49:339–353.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
