

Genome-wide diversity and selective pressure in the human rhinovirus
- PMID: 17477878
- PMCID: PMC1892812
- DOI: 10.1186/1743-422X-4-40
Genome-wide diversity and selective pressure in the human rhinovirus
Abstract
Background: The human rhinoviruses (HRV) are one of the most common and diverse respiratory pathogens of humans. Over 100 distinct HRV serotypes are known, yet only 6 genomes are available. Due to the paucity of HRV genome sequence, little is known about the genetic diversity within HRV or the forces driving this diversity. Previous comparative genome sequence analyses indicate that recombination drives diversification in multiple genera of the picornavirus family, yet it remains unclear if this holds for HRV.
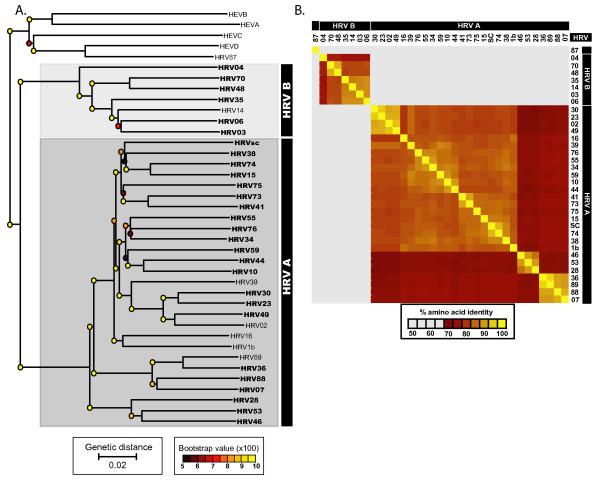
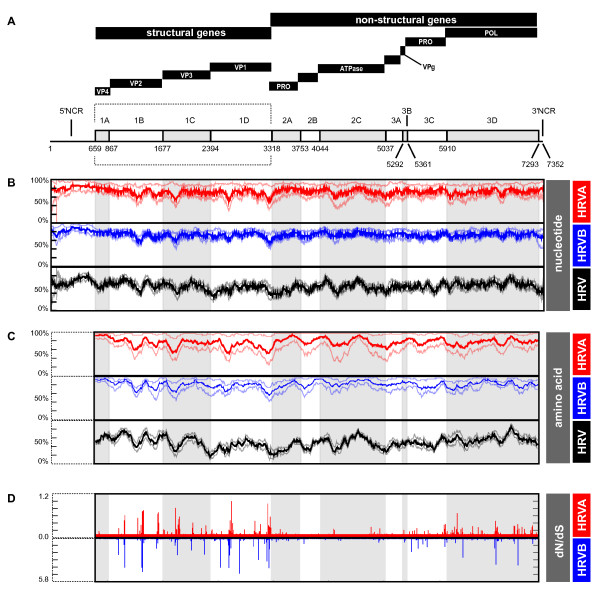
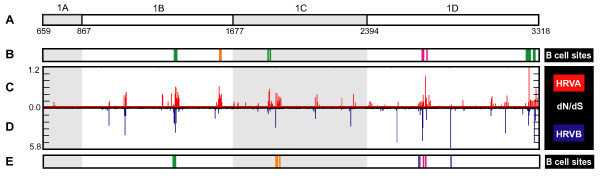
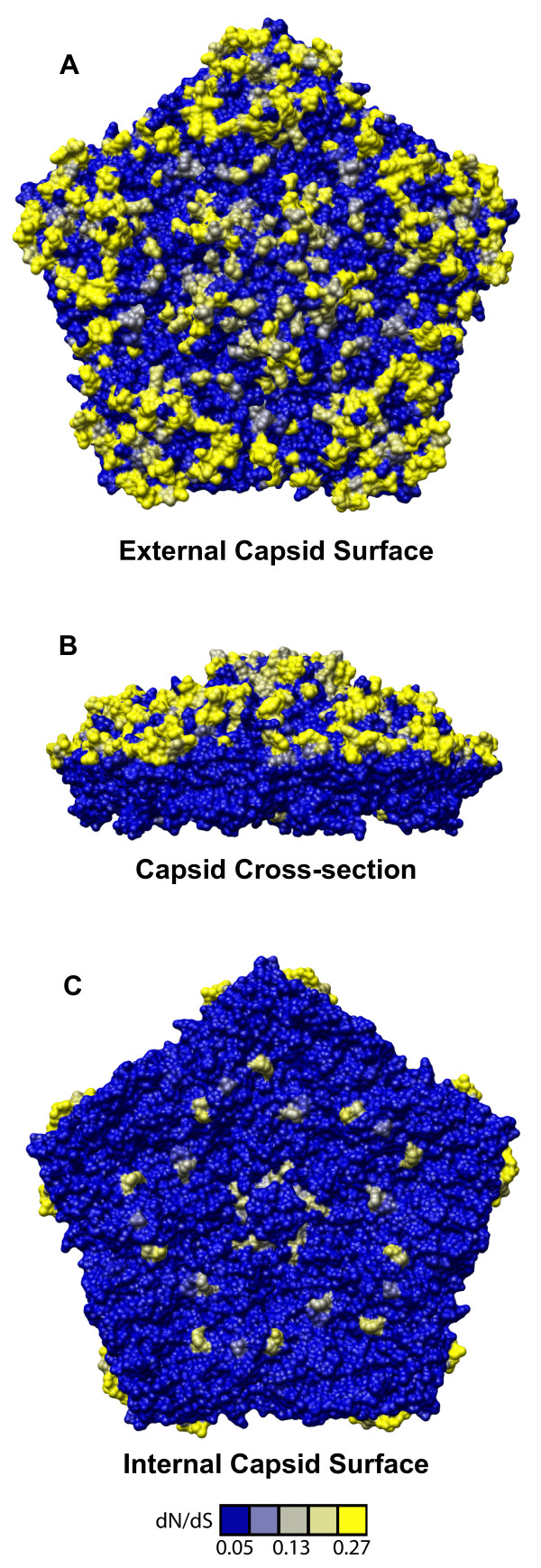
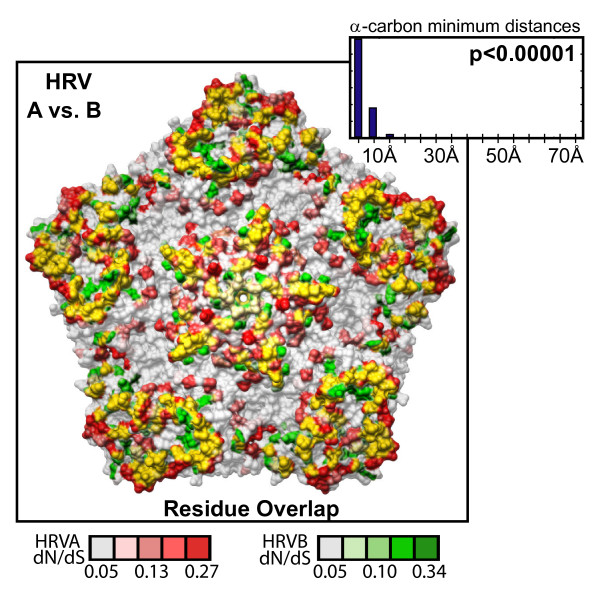
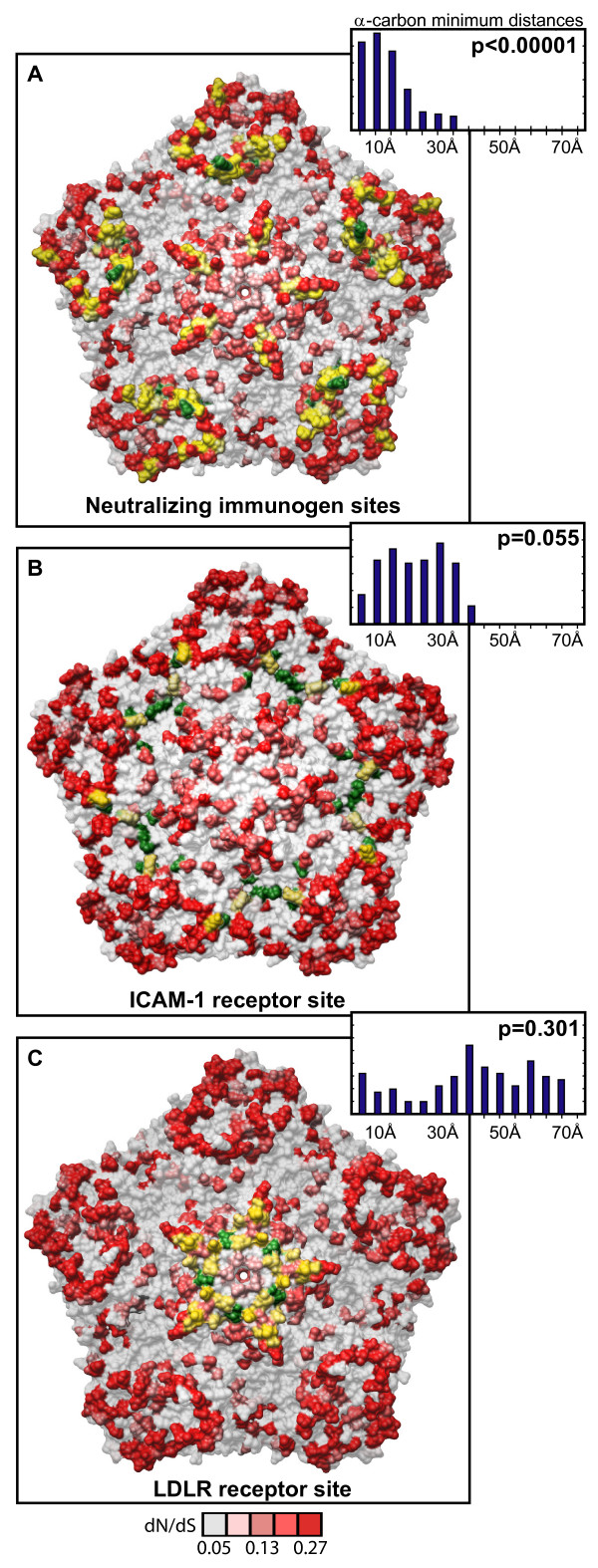
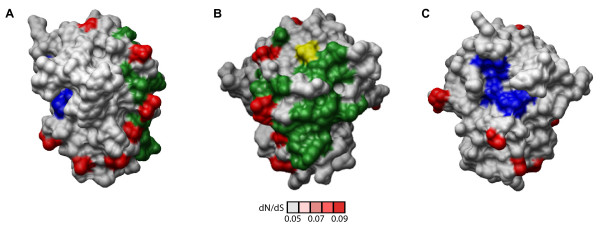
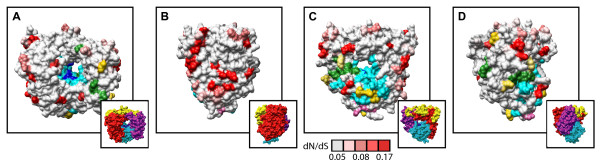
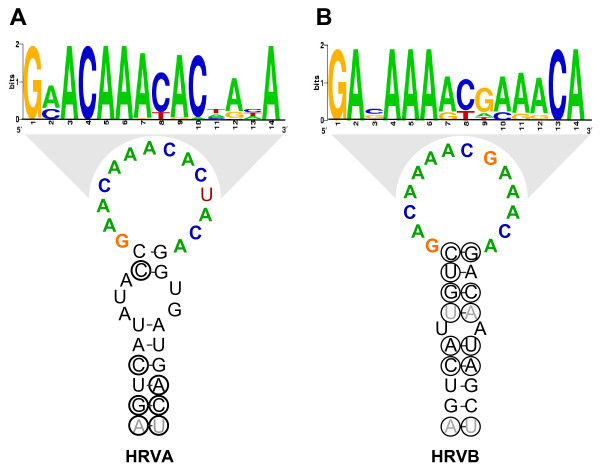
Results: To resolve this and gain insight into the forces driving diversification in HRV, we generated a representative set of 34 fully sequenced HRVs. Analysis of these genomes shows consistent phylogenies across the genome, conserved non-coding elements, and only limited recombination. However, spikes of genetic diversity at both the nucleotide and amino acid level are detectable within every locus of the genome. Despite this, the HRV genome as a whole is under purifying selective pressure, with islands of diversifying pressure in the VP1, VP2, and VP3 structural genes and two non-structural genes, the 3C protease and 3D polymerase. Mapping diversifying residues in these factors onto available 3-dimensional structures revealed the diversifying capsid residues partition to the external surface of the viral particle in statistically significant proximity to antigenic sites. Diversifying pressure in the pleconaril binding site is confined to a single residue known to confer drug resistance (VP1 191). In contrast, diversifying pressure in the non-structural genes is less clear, mapping both nearby and beyond characterized functional domains of these factors.
Conclusion: This work provides a foundation for understanding HRV genetic diversity and insight into the underlying biology driving evolution in HRV. It expands our knowledge of the genome sequence space that HRV reference serotypes occupy and how the pattern of genetic diversity across HRV genomes differs from other picornaviruses. It also reveals evidence of diversifying selective pressure in both structural genes known to interact with the host immune system and in domains of unassigned function in the non-structural 3C and 3D genes, raising the possibility that diversification of undiscovered functions in these essential factors may influence HRV fitness and evolution.
Figures
References
-
- Gern JE, Galagan DM, Jarjour NN, Dick EC, Busse WW. Detection of rhinovirus RNA in lower airway cells during experimentally induced infection. Am J Respir Crit Care Med. 1997;155:1159–1161. - PubMed
-
- Papadopoulos NG, Johnston SL. Rhinoviruses as pathogens of the lower respiratory tract. Can Respir J. 2000;7:409–414. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
