

Intercellular coupling confers robustness against mutations in the SCN circadian clock network
- PMID: 17482552
- PMCID: PMC3749832
- DOI: 10.1016/j.cell.2007.02.047
Intercellular coupling confers robustness against mutations in the SCN circadian clock network
Abstract
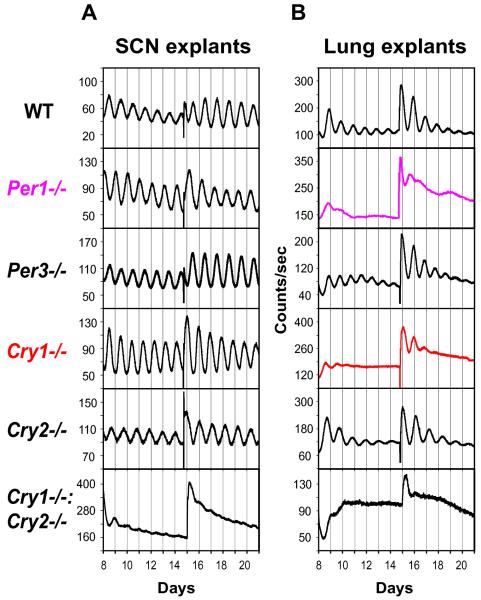
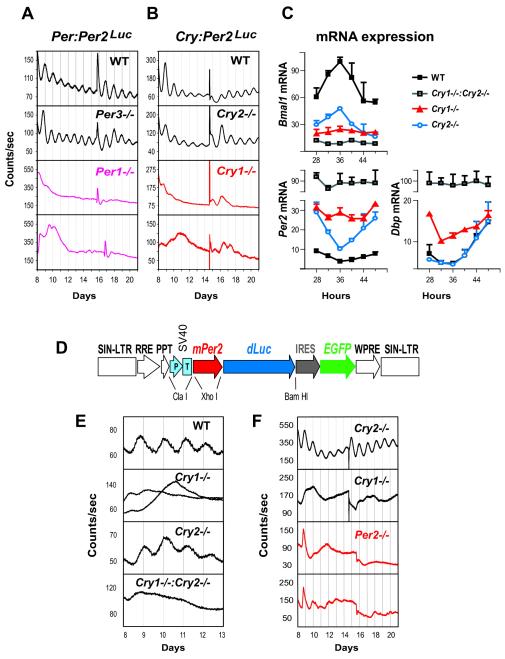
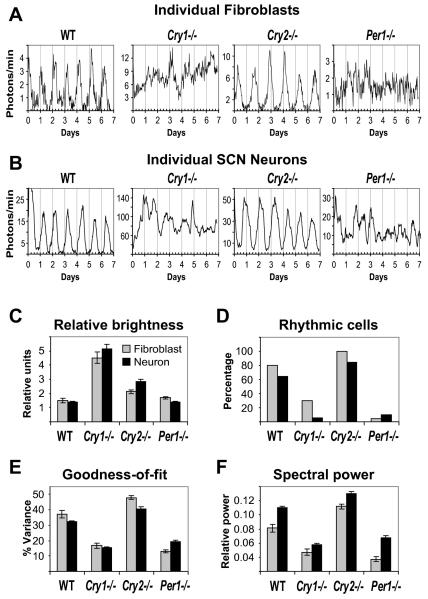
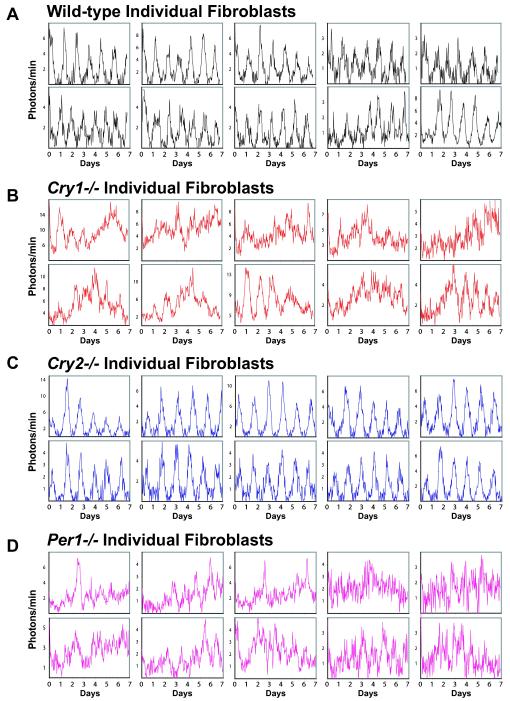
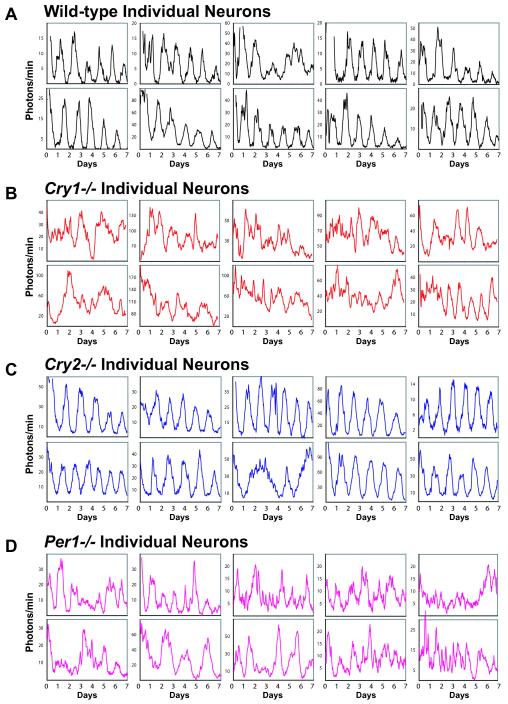
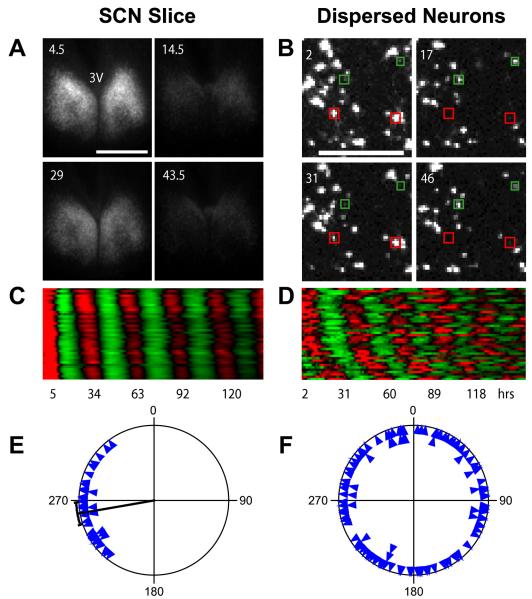
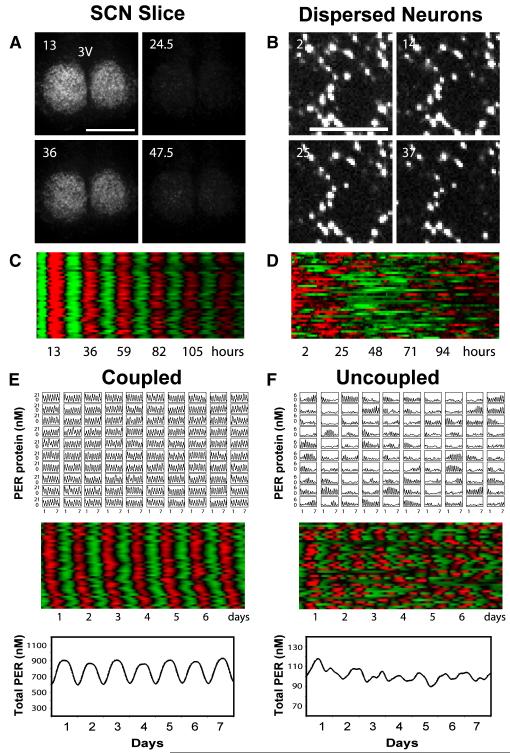
Molecular mechanisms of the mammalian circadian clock have been studied primarily by genetic perturbation and behavioral analysis. Here, we used bioluminescence imaging to monitor Per2 gene expression in tissues and cells from clock mutant mice. We discovered that Per1 and Cry1 are required for sustained rhythms in peripheral tissues and cells, and in neurons dissociated from the suprachiasmatic nuclei (SCN). Per2 is also required for sustained rhythms, whereas Cry2 and Per3 deficiencies cause only period length defects. However, oscillator network interactions in the SCN can compensate for Per1 or Cry1 deficiency, preserving sustained rhythmicity in mutant SCN slices and behavior. Thus, behavior does not necessarily reflect cell-autonomous clock phenotypes. Our studies reveal previously unappreciated requirements for Per1, Per2, and Cry1 in sustaining cellular circadian rhythmicity and demonstrate that SCN intercellular coupling is essential not only to synchronize component cellular oscillators but also for robustness against genetic perturbations.
Figures
References
-
- Bae K, Jin XW, Maywood ES, Hastings MH, Reppert SM, Weaver DR. Differential functions of mPer1, mPer2, and mPer3 in the SCN circadian clock Neuron. 2001;30:525–536. - PubMed
-
- Balsalobre A, Damiola F, Schibler U. A serum shock induces circadian gene expression in mammalian tissue culture cells. Cell. 1998;93:929–937. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
