

Phosphorylation of the Sic1 inhibitor of B-type cyclins in Saccharomyces cerevisiae is not essential but contributes to cell cycle robustness
- PMID: 17483408
- PMCID: PMC1931548
- DOI: 10.1534/genetics.107.073494
Phosphorylation of the Sic1 inhibitor of B-type cyclins in Saccharomyces cerevisiae is not essential but contributes to cell cycle robustness
Abstract
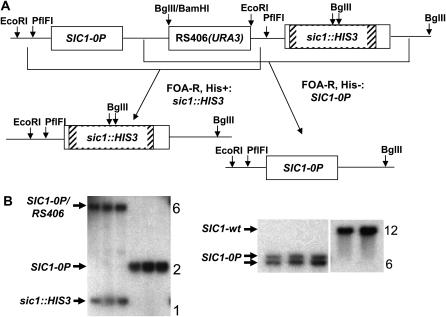
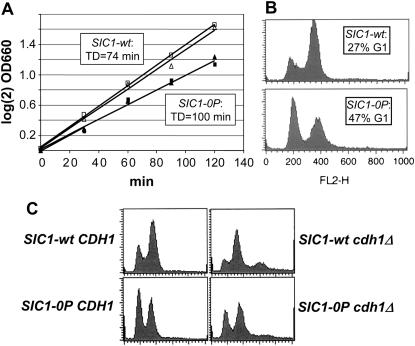
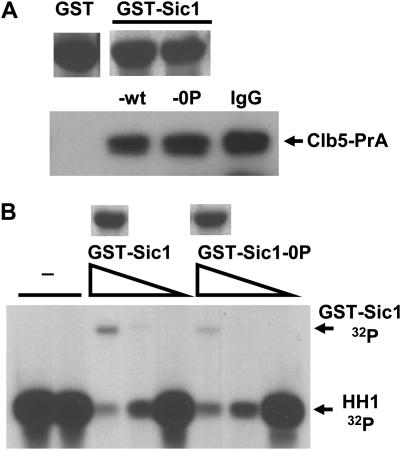
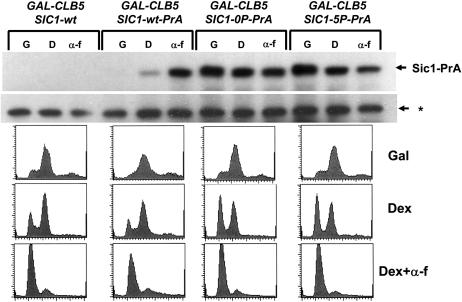
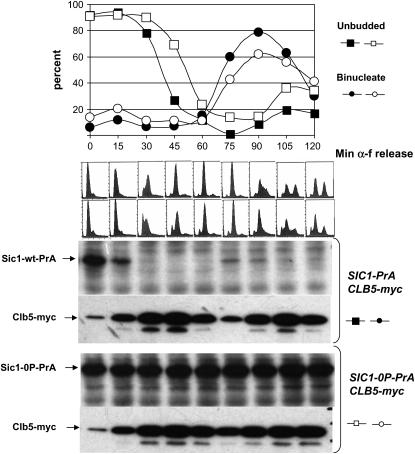
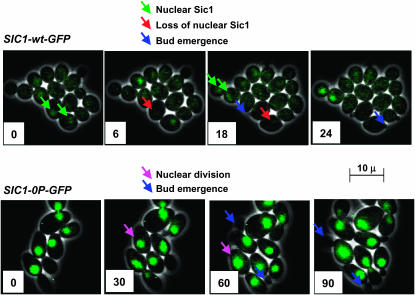
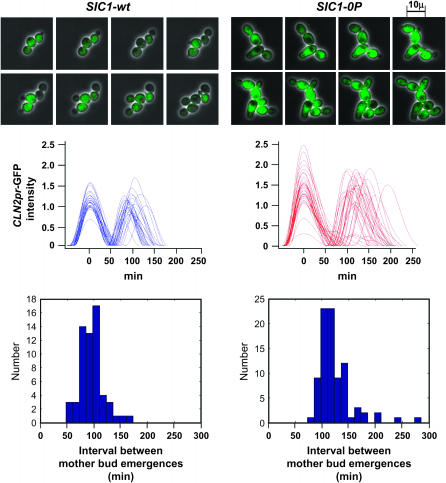
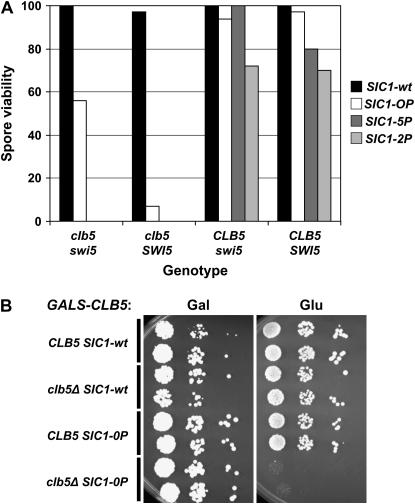
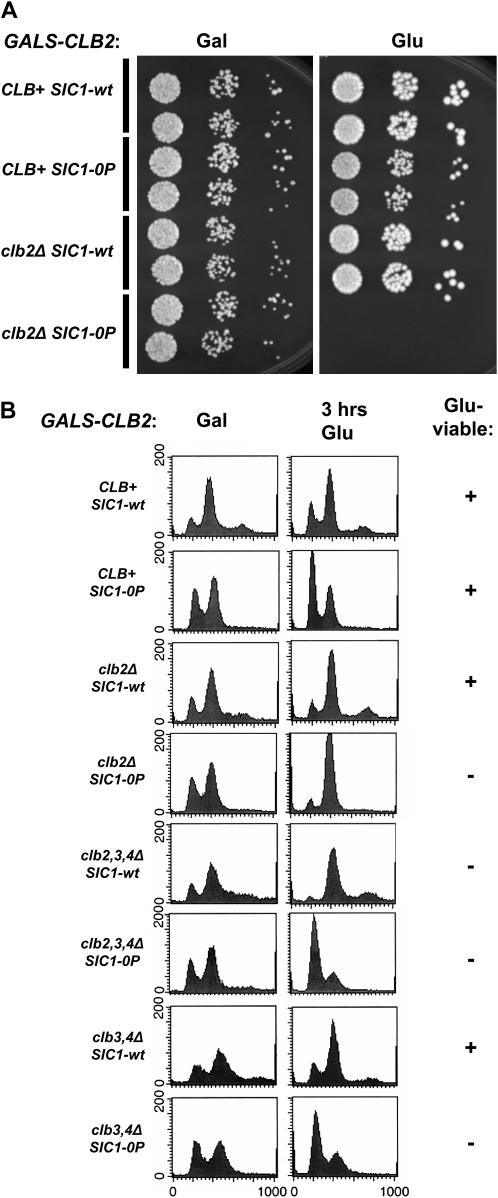
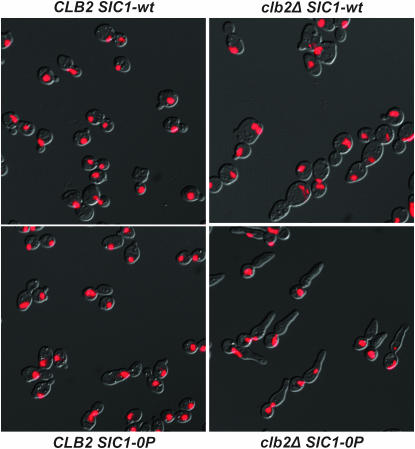
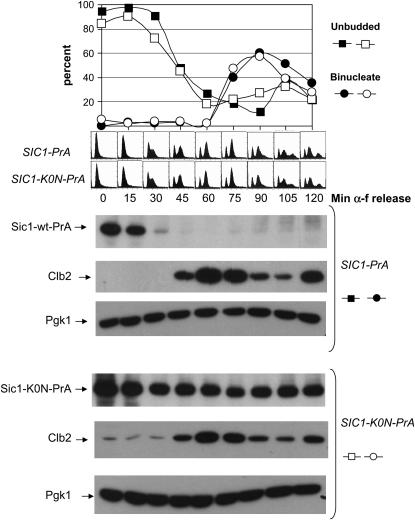
In budding yeast, B-type cyclin (Clb)-dependent kinase activity is essential for S phase and mitosis. In newborn G(1) cells, Clb kinase accumulation is blocked, in part because of the Sic1 stoichiometric inhibitor. Previous results strongly suggested that G(1) cyclin-dependent Sic1 phosphorylation, and its consequent degradation, is essential for S phase. However, cells containing a precise endogenous gene replacement of SIC1 with SIC1-0P (all nine phosphorylation sites mutated) were fully viable. Unphosphorylatable Sic1 was abundant and nuclear throughout the cell cycle and effectively inhibited Clb kinase in vitro. SIC1-0P cells had a lengthened G(1) and increased G(1) cyclin transcriptional activation and variable delays in the budded part of the cell cycle. SIC1-0P was lethal when combined with deletion of CLB2, CLB3, or CLB5, the major B-type cyclins. Sic1 phosphorylation provides a sharp link between G(1) cyclin activation and Clb kinase activation, but failure of Sic1 phosphorylation and proteolysis imposes a variable cell cycle delay and extreme sensitivity to B-type cyclin dosage, rather than a lethal cell cycle block.
Figures
References
-
- Amon, A., M. Tyers, B. Futcher and K. Nasmyth, 1993. Mechanisms that help the yeast cell cycle clock tick: G2 cyclins transcriptionally activate G2 cyclins and repress G1 cyclins. Cell 74: 993–1007. - PubMed
-
- Bar-Even, A., J. Paulsson, N. Maheshri, M. Carmi, E. O'Shea et al., 2006. Noise in protein expression scales with natural protein abundance. Nat. Genet. 38: 636–643. - PubMed
-
- Bean, J. M., E. D. Siggia and F. R. Cross, 2006. Coherence and timing of cell cycle Start examined at single-cell resolution. Mol. Cell 21: 3–14. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
