

Oxidative stress enzymes are required for DAF-16-mediated immunity due to generation of reactive oxygen species by Caenorhabditis elegans
- PMID: 17483415
- PMCID: PMC1931534
- DOI: 10.1534/genetics.107.072587
Oxidative stress enzymes are required for DAF-16-mediated immunity due to generation of reactive oxygen species by Caenorhabditis elegans
Abstract
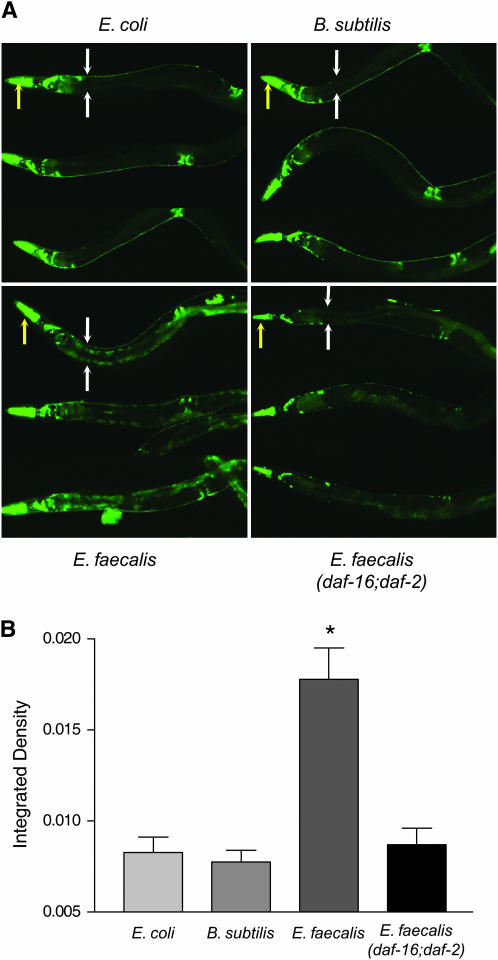
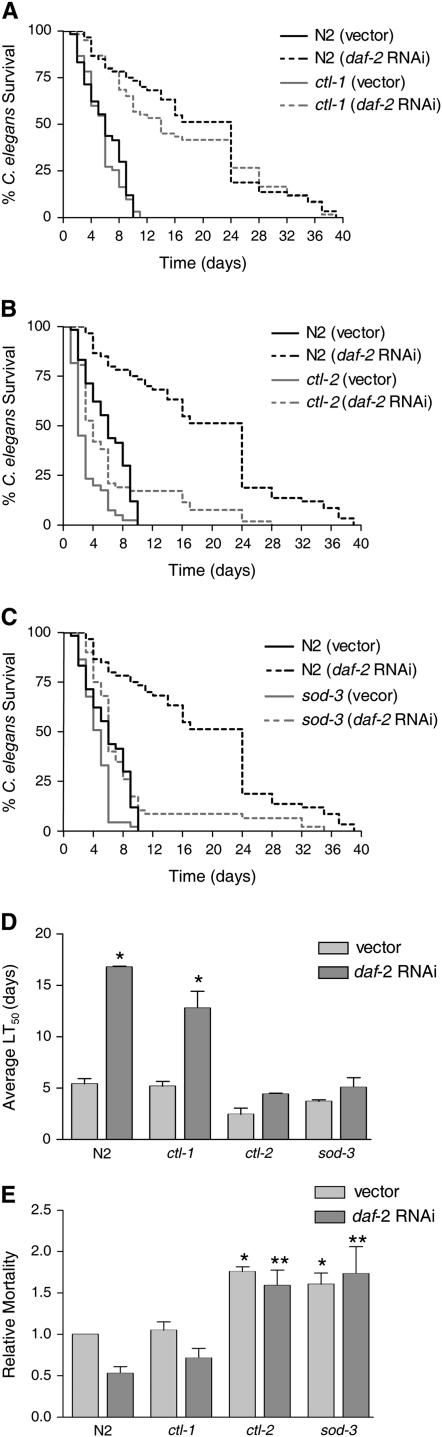
Caenorhabditis elegans has recently been developed as a model for microbial pathogenesis, yet little is known about its immunological defenses. Previous work implicated insulin signaling in mediating pathogen resistance in a manner dependent on the transcriptional regulator DAF-16, but the mechanism has not been elucidated. We present evidence that C. elegans, like mammalian phagocytes, produces reactive oxygen species (ROS) in response to pathogens. Signs of oxidative stress occur in the intestine - the site of the host-pathogen interface - suggesting that ROS release is localized to this tissue. Evidence includes the accumulation of lipofuscin, a pigment resulting from oxidative damage, at this site. In addition, SOD-3, a superoxide dismutase regulated by DAF-16, is induced in intestinal tissue after exposure to pathogenic bacteria. Moreover, we show that the oxidative stress response genes sod-3 and ctl-2 are required for DAF-16-mediated resistance to Enterococcus faecalis using a C. elegans killing assay. We propose a model whereby C. elegans responds to pathogens by producing ROS in the intestine while simultaneously inducing a DAF-16-dependent oxidative stress response to protect adjacent tissues. Because insulin-signaling mutants overproduce oxidative stress response enzymes, the model provides an explanation for their increased resistance to pathogens.
Figures





References
-
- Apel, K., and H. Hirt, 2004. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 55: 373–399. - PubMed
-
- Balaban, R. S., S. Nemoto and T. Finkel, 2005. Mitochondria, oxidants, and aging. Cell 120: 483–495. - PubMed
-
- Bolm, M., G. S. Chhatwal and W. T. Jansen, 2004. a Bacterial resistance of daf-2 mutants. Science 303: 1976; author reply 1976. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
