

Microhomology-mediated end joining in fission yeast is repressed by pku70 and relies on genes involved in homologous recombination
- PMID: 17483423
- PMCID: PMC1931558
- DOI: 10.1534/genetics.107.071621
Microhomology-mediated end joining in fission yeast is repressed by pku70 and relies on genes involved in homologous recombination
Abstract
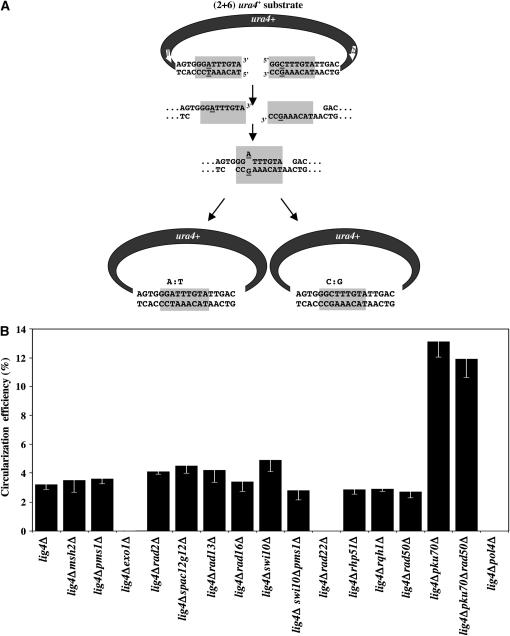
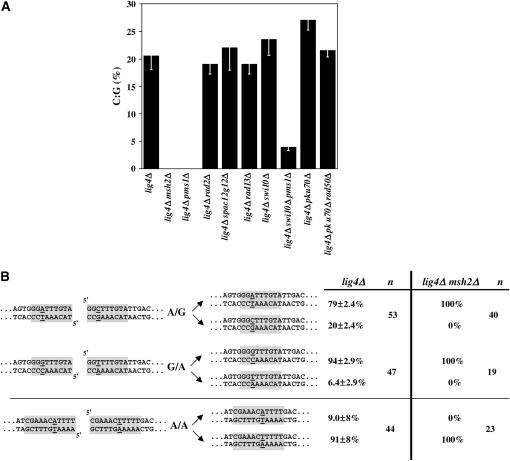
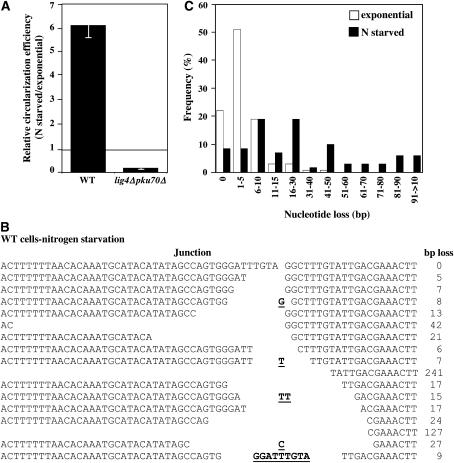
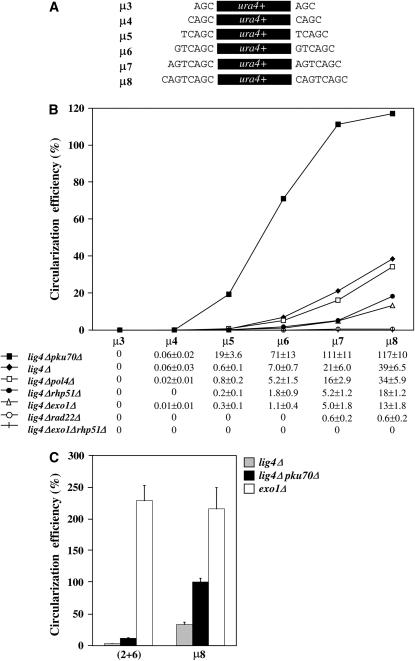
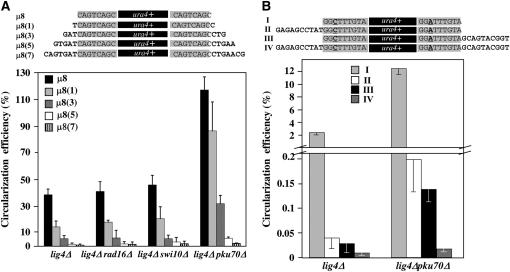
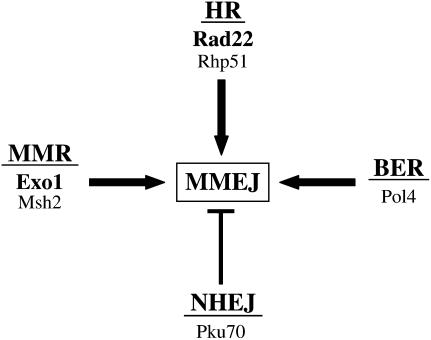
Two DNA repair pathways are known to mediate DNA double-strand-break (DSB) repair: homologous recombination (HR) and nonhomologous end joining (NHEJ). In addition, a nonconservative backup pathway showing extensive nucleotide loss and relying on microhomologies at repair junctions was identified in NHEJ-deficient cells from a variety of organisms and found to be involved in chromosomal translocations. Here, an extrachromosomal assay was used to characterize this microhomology-mediated end-joining (MMEJ) mechanism in fission yeast. MMEJ was found to require at least five homologous nucleotides and its efficiency was decreased by the presence of nonhomologous nucleotides either within the overlapping sequences or at DSB ends. Exo1 exonuclease and Rad22, a Rad52 homolog, were required for repair, suggesting that MMEJ is related to the single-strand-annealing (SSA) pathway of HR. In addition, MMEJ-dependent repair of DSBs with discontinuous microhomologies was strictly dependent on Pol4, a PolX DNA polymerase. Although not strictly required, Msh2 and Pms1 mismatch repair proteins affected the pattern of MMEJ repair. Strikingly, Pku70 inhibited MMEJ and increased the minimal homology length required for efficient MMEJ. Overall, this study strongly suggests that MMEJ does not define a distinct DSB repair mechanism but reflects "micro-SSA."
Figures
Similar articles
-
Assays for DNA double-strand break repair by microhomology-based end-joining repair mechanisms.Nucleic Acids Res. 2016 Apr 7;44(6):e56. doi: 10.1093/nar/gkv1349. Epub 2015 Dec 10. Nucleic Acids Res. 2016. PMID: 26657630 Free PMC article.
-
Microhomology-mediated End Joining and Homologous Recombination share the initial end resection step to repair DNA double-strand breaks in mammalian cells.Proc Natl Acad Sci U S A. 2013 May 7;110(19):7720-5. doi: 10.1073/pnas.1213431110. Epub 2013 Apr 22. Proc Natl Acad Sci U S A. 2013. PMID: 23610439 Free PMC article.
-
Single-Strand Annealing Plays a Major Role in Double-Strand DNA Break Repair following CRISPR-Cas9 Cleavage in Leishmania.mSphere. 2019 Aug 21;4(4):e00408-19. doi: 10.1128/mSphere.00408-19. mSphere. 2019. PMID: 31434745 Free PMC article.
-
Risky business: Microhomology-mediated end joining.Mutat Res. 2016 Jun;788:17-24. doi: 10.1016/j.mrfmmm.2015.12.005. Epub 2016 Jan 2. Mutat Res. 2016. PMID: 26790771 Free PMC article. Review.
-
Contribution of Microhomology to Genome Instability: Connection between DNA Repair and Replication Stress.Int J Mol Sci. 2022 Oct 26;23(21):12937. doi: 10.3390/ijms232112937. Int J Mol Sci. 2022. PMID: 36361724 Free PMC article. Review.
Cited by
-
Microhomology-mediated deletion and gene conversion in African trypanosomes.Nucleic Acids Res. 2011 Mar;39(4):1372-80. doi: 10.1093/nar/gkq981. Epub 2010 Oct 21. Nucleic Acids Res. 2011. PMID: 20965968 Free PMC article.
-
Advancing metabolic engineering of Yarrowia lipolytica using the CRISPR/Cas system.Appl Microbiol Biotechnol. 2018 Nov;102(22):9541-9548. doi: 10.1007/s00253-018-9366-x. Epub 2018 Sep 21. Appl Microbiol Biotechnol. 2018. PMID: 30238143 Free PMC article. Review.
-
Is non-homologous end-joining really an inherently error-prone process?PLoS Genet. 2014 Jan;10(1):e1004086. doi: 10.1371/journal.pgen.1004086. Epub 2014 Jan 16. PLoS Genet. 2014. PMID: 24453986 Free PMC article. Review.
-
Microhomology-mediated end joining is activated in irradiated human cells due to phosphorylation-dependent formation of the XRCC1 repair complex.Nucleic Acids Res. 2017 Mar 17;45(5):2585-2599. doi: 10.1093/nar/gkw1262. Nucleic Acids Res. 2017. PMID: 27994036 Free PMC article.
-
The helicase domain of Polθ counteracts RPA to promote alt-NHEJ.Nat Struct Mol Biol. 2017 Dec;24(12):1116-1123. doi: 10.1038/nsmb.3494. Epub 2017 Oct 23. Nat Struct Mol Biol. 2017. PMID: 29058711 Free PMC article.
References
-
- Bahler, J., J. Q. Wu, M. S. Longtine, N. G. Shah, A. McKenzie, III et al., 1998. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast 14: 943–951. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
