

Decondensing the protamine domain for transcription
- PMID: 17483471
- PMCID: PMC1895951
- DOI: 10.1073/pnas.0700076104
Decondensing the protamine domain for transcription
Abstract
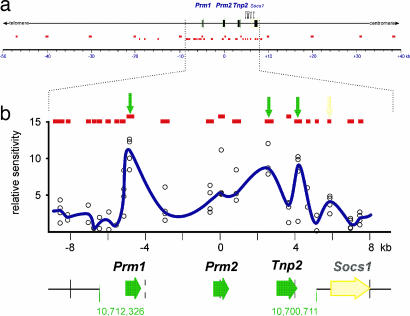
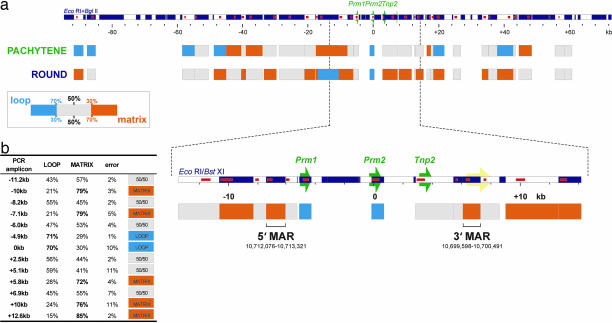
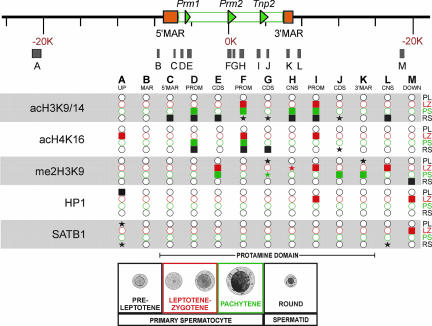
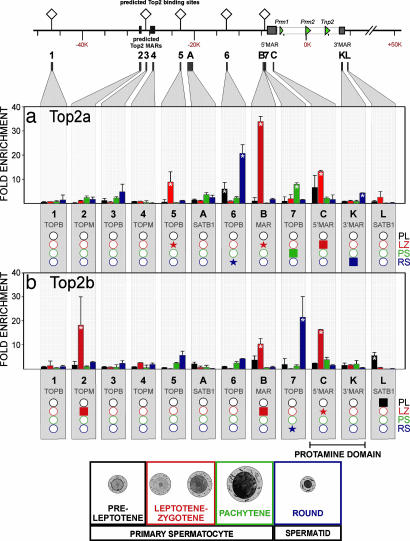
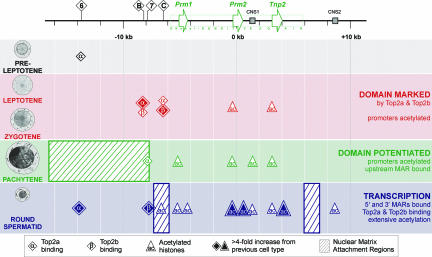
Potentiation is the transition from higher-order, transcriptionally silent chromatin to a less condensed state requisite to accommodating the molecular elements required for transcription. To examine the underlying mechanism of potentiation an approximately 13.7-kb mouse protamine domain of increased nuclease sensitivity flanked by 5' and 3' nuclear matrix attachment regions was defined. The potentiated DNase I-sensitive region is formed at the pachytene spermatocyte stage with the recruitment to the nuclear matrix of a large approximately 9.6-kb region just upstream of the domain. Attachment is then specified in the transcribing round spermatid, recapitulating the organization of the human cluster. In comparison to other modifiers that have no effect, i.e., histone methylation, HP1, and SATB1, topoisomerase engages nuclear matrix binding as minor marks of histone acetylation appear. Reorganization is marked by specific sites of topoisomerase II activity that are initially detected in leptotene-zygotene spermatocytes just preceding the formation of the DNase I-sensitive domain. This has provided a likely model of the events initiating potentiation, i.e., the opening of a chromatin domain.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Choudhary SK, Wykes SM, Kramer JA, Mohamed AN, Koppitch F, Nelson JE, Krawetz SA. J Biol Chem. 1995;270:8755–8762. - PubMed
-
- Chambeyron S, Da Silva NR, Lawson KA, Bickmore WA. Development (Cambridge, UK) 2005;132:2215–2223. - PubMed
-
- Keene MA, Corces V, Lowenhaupt K, Elgin SCR. Eur J Cell Biol. 1980;22:95–95.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
