

Lysosomal integral membrane protein 2 is a novel component of the cardiac intercalated disc and vital for load-induced cardiac myocyte hypertrophy
- PMID: 17485520
- PMCID: PMC2118572
- DOI: 10.1084/jem.20070145
Lysosomal integral membrane protein 2 is a novel component of the cardiac intercalated disc and vital for load-induced cardiac myocyte hypertrophy
Abstract
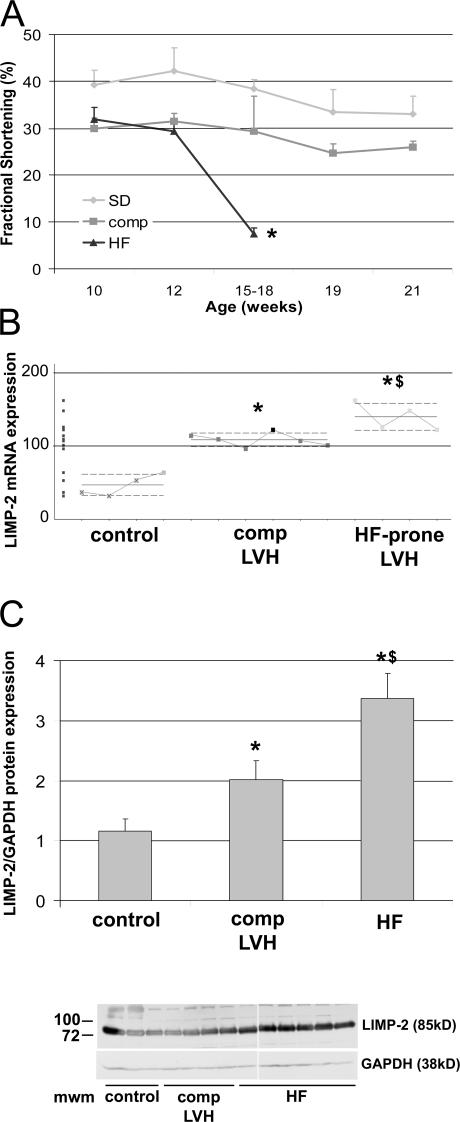
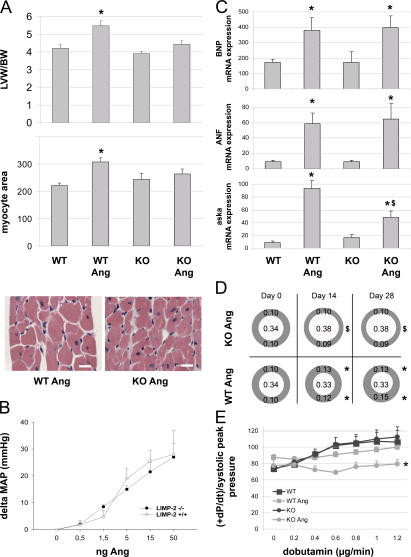
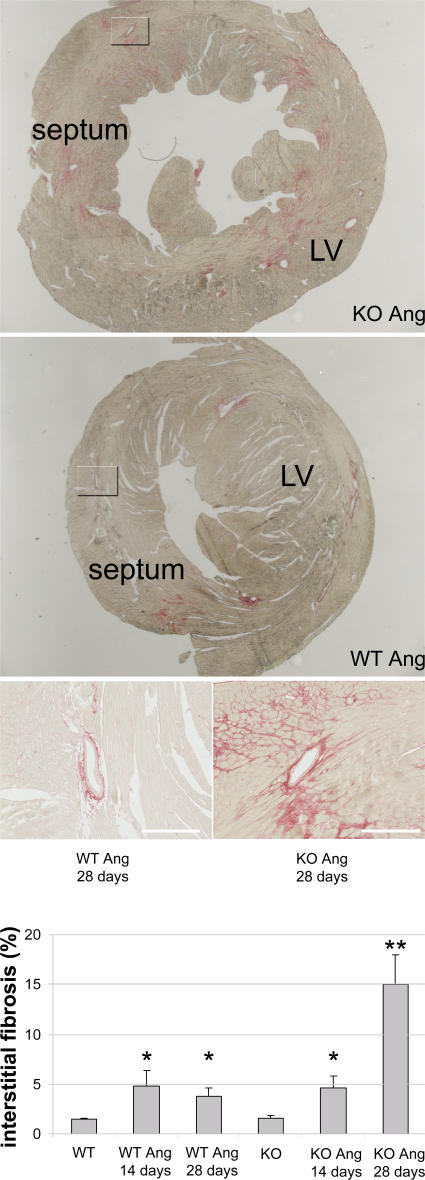
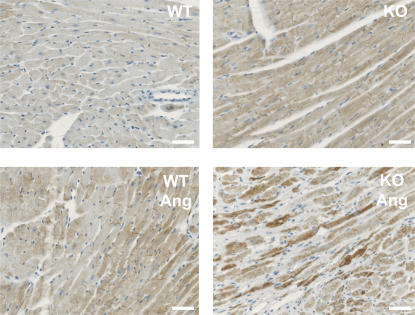
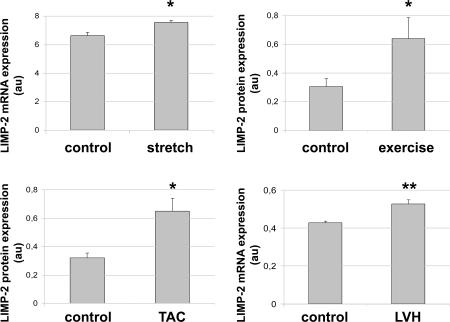
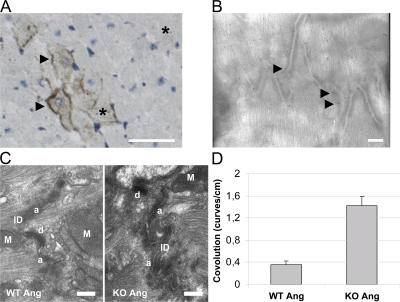
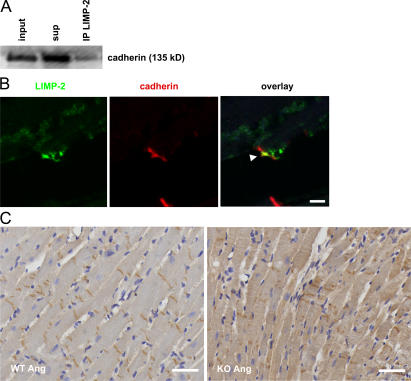
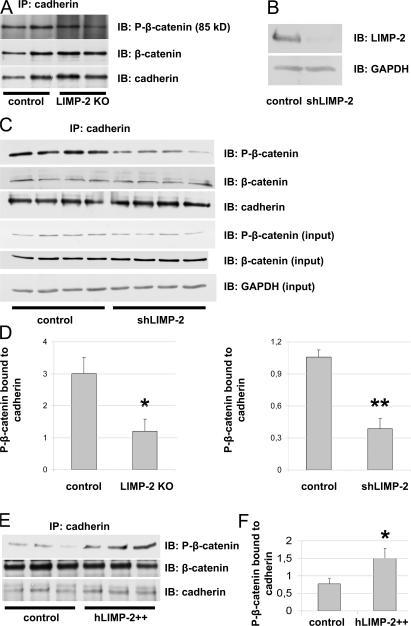
The intercalated disc (ID) of cardiac myocytes is emerging as a crucial structure in the heart. Loss of ID proteins like N-cadherin causes lethal cardiac abnormalities, and mutations in ID proteins cause human cardiomyopathy. A comprehensive screen for novel mechanisms in failing hearts demonstrated that expression of the lysosomal integral membrane protein 2 (LIMP-2) is increased in cardiac hypertrophy and heart failure in both rat and human myocardium. Complete loss of LIMP-2 in genetically engineered mice did not affect cardiac development; however, these LIMP-2 null mice failed to mount a hypertrophic response to increased blood pressure but developed cardiomyopathy. Disturbed cadherin localization in these hearts suggested that LIMP-2 has important functions outside lysosomes. Indeed, we also find LIMP-2 in the ID, where it associates with cadherin. RNAi-mediated knockdown of LIMP-2 decreases the binding of phosphorylated beta-catenin to cadherin, whereas overexpression of LIMP-2 has the opposite effect. Collectively, our data show that LIMP-2 is crucial to mount the adaptive hypertrophic response to cardiac loading. We demonstrate a novel role for LIMP-2 as an important mediator of the ID.
Figures
References
-
- Opie, L.H. 2006. Controversies in cardiology. Lancet. 367:13–14. - PubMed
-
- Perriard, J.C., A. Hirschy, and E. Ehler. 2003. Dilated cardiomyopathy: a disease of the intercalated disc? Trends Cardiovasc. Med. 13:30–38. - PubMed
-
- Radice, G.L., H. Rayburn, H. Matsunami, K.A. Knudsen, M. Takeichi, and R.O. Hynes. 1997. Developmental defects in mouse embryos lacking N-cadherin. Dev. Biol. 181:64–78. - PubMed
-
- Haegel, H., L. Larue, M. Ohsugi, L. Fedorov, K. Herrenknecht, and R. Kemler. 1995. Lack of beta-catenin affects mouse development at gastrulation. Development. 121:3529–3537. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
