

Reprogramming the tRNA-splicing activity of a bacterial RNA repair enzyme
- PMID: 17488852
- PMCID: PMC1920235
- DOI: 10.1093/nar/gkm110
Reprogramming the tRNA-splicing activity of a bacterial RNA repair enzyme
Abstract
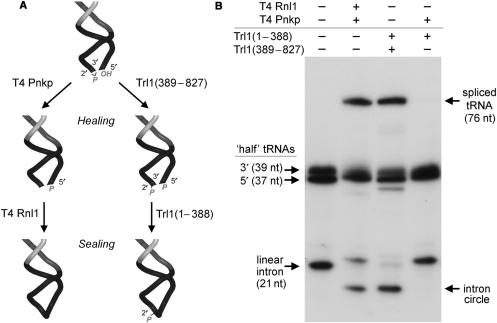
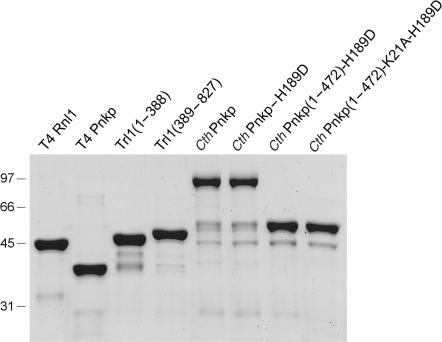
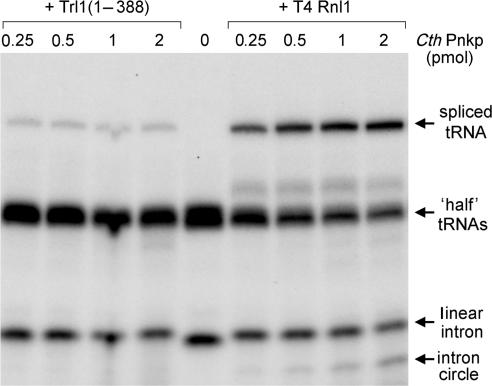
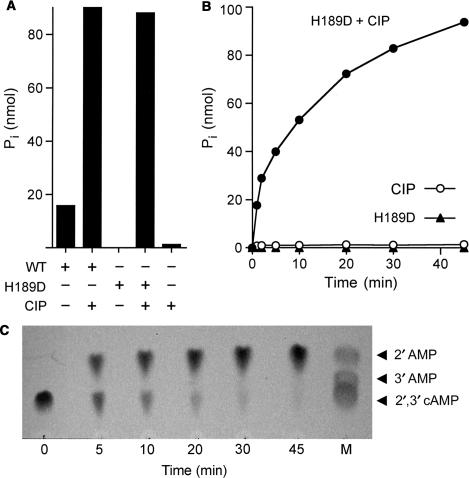
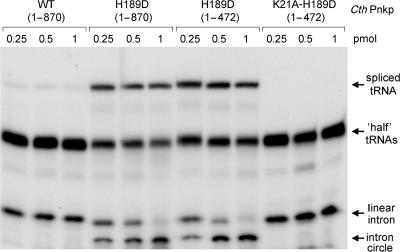
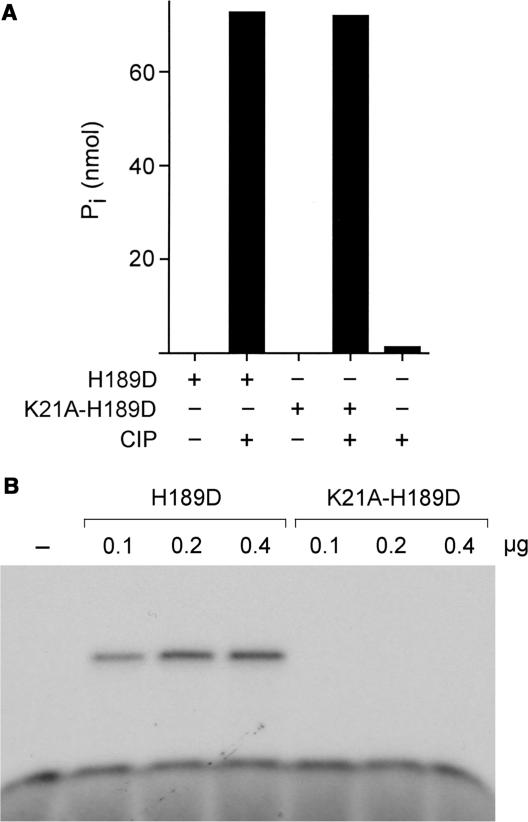
Programmed RNA breakage is an emerging theme underlying cellular responses to stress, virus infection and defense against foreign species. In many cases, site-specific cleavage of the target RNA generates 2',3' cyclic phosphate and 5'-OH ends. For the damage to be repaired, both broken ends must be healed before they can be sealed by a ligase. Healing entails hydrolysis of the 2',3' cyclic phosphate to form a 3'-OH and phosphorylation of the 5'-OH to form a 5'-PO4. Here, we demonstrate that a polynucleotide kinase-phosphatase enzyme from Clostridium thermocellum (CthPnkp) can catalyze both of the end-healing steps of tRNA splicing in vitro. The route of tRNA repair by CthPnkp can be reprogrammed by a mutation in the 3' end-healing domain (H189D) that yields a 2'-PO4 product instead of a 2'-OH. Whereas tRNA ends healed by wild-type CthPnkp are readily sealed by T4 RNA ligase 1, the H189D enzyme generates ends that are spliced by yeast tRNA ligase. Our findings suggest that RNA repair enzymes can evolve their specificities to suit a particular pathway.
Figures
References
-
- Abelson J, Trotta CR, Li H. tRNA splicing. J. Biol. Chem. 1998;273:12685–12688. - PubMed
-
- Sidrauski C, Cox JS, Walter P. tRNA ligase is required for regulated mRNA splicing in the unfolded protein response. Cell. 1996;87:405–413. - PubMed
-
- Greer CL, Peebles CL, Gegenheimer P, Abelson J. Mechanism of action of a yeast RNA ligase in tRNA splicing. Cell. 1983;32:537–546. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
