

Mice lacking the signaling molecule CalDAG-GEFI represent a model for leukocyte adhesion deficiency type III
- PMID: 17492052
- PMCID: PMC1865026
- DOI: 10.1172/JCI30575
Mice lacking the signaling molecule CalDAG-GEFI represent a model for leukocyte adhesion deficiency type III
Abstract
Single gene mutations in beta integrins can account for functional defects of individual cells of the hematopoietic system. In humans, mutations in beta(2) integrin lead to leukocyte adhesion deficiency (LAD) syndrome and mutations in beta(3) integrin cause the bleeding disorder Glanzmann thrombasthenia. However, multiple defects in blood cells involving various beta integrins (beta(1), beta(2), and beta(3)) occur simultaneously in patients with the recently described LAD type III (LAD-III). Here we show that the product of a single gene, Ca(2+) and diacylglycerol-regulated guanine nucleotide exchange factor I (CalDAG-GEFI), controlled the activation of all 3 integrins in the hematopoietic system. Neutrophils from CalDAG-GEFI(-/-) mice exhibited strong defects in Rap1 and beta(1) and beta(2) integrin activation while maintaining normal calcium flux, degranulation, and ROS generation. Neutrophils from CalDAG-GEFI-deficient mice failed to adhere firmly to stimulated venules and to migrate into sites of inflammation. Furthermore, CalDAG-GEFI regulated the activation of beta(1) and beta(3) integrins in platelets, and CalDAG-GEFI deficiency caused complete inhibition of arterial thrombus formation in mice. Thus, mice engineered to lack CalDAG-GEFI have a combination of defects in leukocyte and platelet functions similar to that of LAD-III patients.
Figures
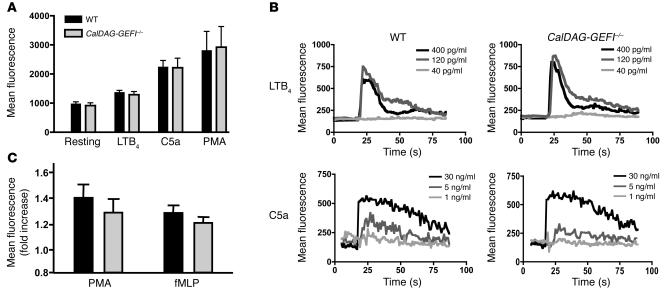


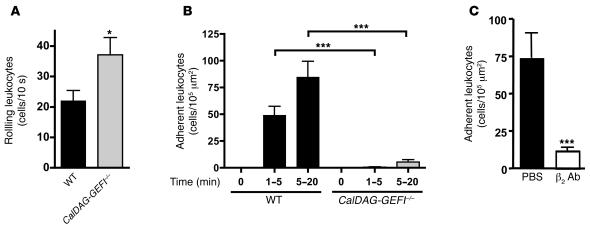
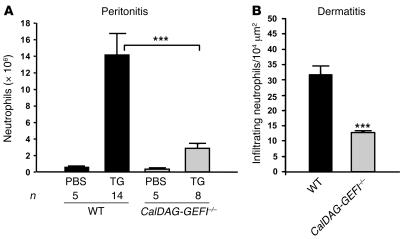

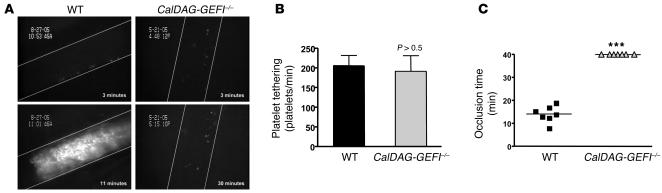
References
-
- Hynes R.O. Integrins: bidirectional, allosteric signaling machines. Cell. 2002;110:673–687. - PubMed
-
- Sims T.N., Dustin M.L. The immunological synapse: integrins take the stage. Immunol. Rev. 2002;186:100–117. - PubMed
-
- Simon S.I., Green C.E. Molecular mechanics and dynamics of leukocyte recruitment during inflammation. Annu. Rev. Biomed. Eng. 2005;7:151–185. - PubMed
-
- Savage B., Cattaneo M., Ruggeri Z.M. Mechanisms of platelet aggregation. Curr. Opin. Hematol. 2001;8:270–276. - PubMed
-
- Anderson D.C., Springer T.A. Leukocyte adhesion deficiency: an inherited defect in the Mac-1, LFA-1, and p150,95 glycoproteins. Annu. Rev. Med. 1987;38:175–194. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
