

Molecular analysis of the sea anemone toxin Av3 reveals selectivity to insects and demonstrates the heterogeneity of receptor site-3 on voltage-gated Na+ channels
- PMID: 17492942
- PMCID: PMC1948988
- DOI: 10.1042/BJ20070233
Molecular analysis of the sea anemone toxin Av3 reveals selectivity to insects and demonstrates the heterogeneity of receptor site-3 on voltage-gated Na+ channels
Abstract
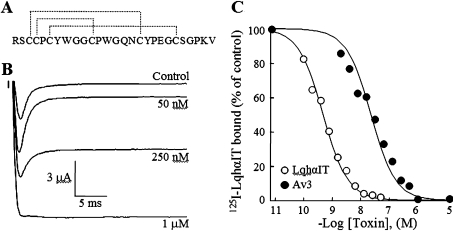
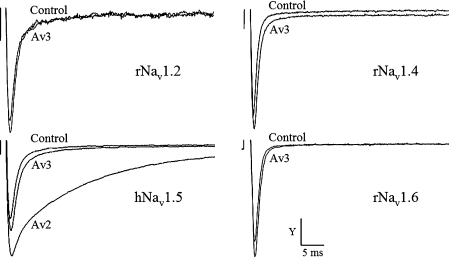
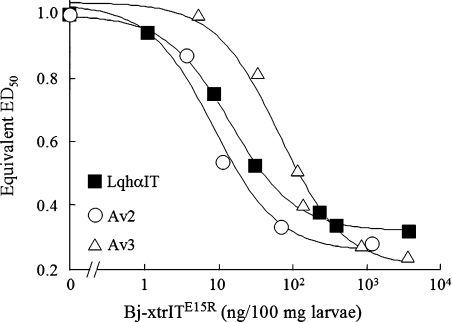
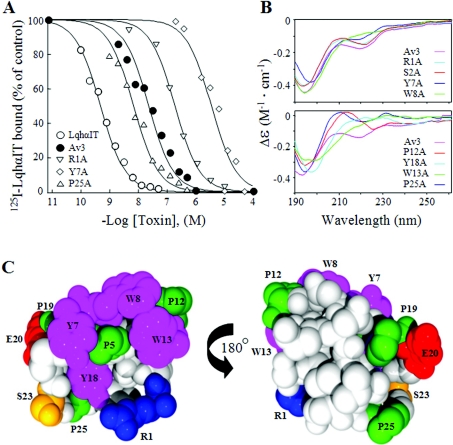
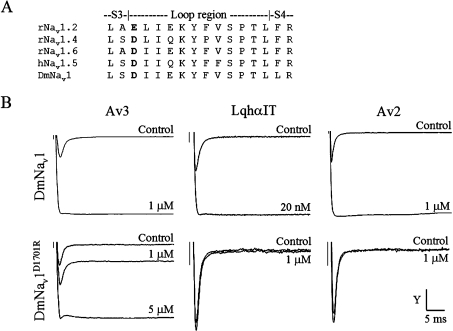
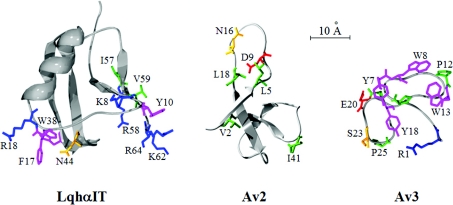
Av3 is a short peptide toxin from the sea anemone Anemonia viridis shown to be active on crustaceans and inactive on mammals. It inhibits inactivation of Na(v)s (voltage-gated Na+ channels) like the structurally dissimilar scorpion alpha-toxins and type I sea anemone toxins that bind to receptor site-3. To examine the potency and mode of interaction of Av3 with insect Na(v)s, we established a system for its expression, mutagenized it throughout, and analysed it in toxicity, binding and electrophysiological assays. The recombinant Av3 was found to be highly toxic to blowfly larvae (ED50=2.65+/-0.46 pmol/100 mg), to compete well with the site-3 toxin LqhalphaIT (from the scorpion Leiurus quinquestriatus) on binding to cockroach neuronal membranes (K(i)=21.4+/-7.1 nM), and to inhibit the inactivation of Drosophila melanogaster channel, DmNa(v)1, but not that of mammalian Na(v)s expressed in Xenopus oocytes. Moreover, like other site-3 toxins, the activity of Av3 was synergically enhanced by ligands of receptor site-4 (e.g. scorpion beta-toxins). The bioactive surface of Av3 was found to consist mainly of aromatic residues and did not resemble any of the bioactive surfaces of other site-3 toxins. These analyses have portrayed a toxin that might interact with receptor site-3 in a different fashion compared with other ligands of this site. This assumption was corroborated by a D1701R mutation in DmNa(v)1, which has been shown to abolish the activity of all other site-3 ligands, except Av3. All in all, the present study provides further evidence for the heterogeneity of receptor site-3, and raises Av3 as a unique model for design of selective anti-insect compounds.
Figures
Similar articles
-
Expression and mutagenesis of the sea anemone toxin Av2 reveals key amino acid residues important for activity on voltage-gated sodium channels.Biochemistry. 2006 Jul 25;45(29):8864-73. doi: 10.1021/bi060386b. Biochemistry. 2006. PMID: 16846229
-
A natural point mutation changes both target selectivity and mechanism of action of sea anemone toxins.FASEB J. 2012 Dec;26(12):5141-51. doi: 10.1096/fj.12-218479. Epub 2012 Sep 12. FASEB J. 2012. PMID: 22972919
-
The specificity of Av3 sea anemone toxin for arthropods is determined at linker DI/SS2-S6 in the pore module of target sodium channels.Biochem J. 2014 Oct 15;463(2):271-7. doi: 10.1042/BJ20140576. Biochem J. 2014. PMID: 25055135
-
Actions of sea anemone type 1 neurotoxins on voltage-gated sodium channel isoforms.Toxicon. 2009 Dec 15;54(8):1102-11. doi: 10.1016/j.toxicon.2009.04.018. Epub 2009 Apr 23. Toxicon. 2009. PMID: 19393679 Review.
-
Sea anemone venom as a source of insecticidal peptides acting on voltage-gated Na+ channels.Toxicon. 2007 Mar 15;49(4):550-60. doi: 10.1016/j.toxicon.2006.11.029. Epub 2006 Dec 5. Toxicon. 2007. PMID: 17224168 Free PMC article. Review.
Cited by
-
Ancient Venom Systems: A Review on Cnidaria Toxins.Toxins (Basel). 2015 Jun 18;7(6):2251-71. doi: 10.3390/toxins7062251. Toxins (Basel). 2015. PMID: 26094698 Free PMC article. Review.
-
Sea Anemone Toxins: A Structural Overview.Mar Drugs. 2019 Jun 1;17(6):325. doi: 10.3390/md17060325. Mar Drugs. 2019. PMID: 31159357 Free PMC article. Review.
-
Charge substitutions at the voltage-sensing module of domain III enhance actions of site-3 and site-4 toxins on an insect sodium channel.Insect Biochem Mol Biol. 2021 Oct;137:103625. doi: 10.1016/j.ibmb.2021.103625. Epub 2021 Aug 3. Insect Biochem Mol Biol. 2021. PMID: 34358664 Free PMC article.
-
Sea anemone (Cnidaria, Anthozoa, Actiniaria) toxins: an overview.Mar Drugs. 2012 Aug;10(8):1812-1851. doi: 10.3390/md10081812. Epub 2012 Aug 22. Mar Drugs. 2012. PMID: 23015776 Free PMC article. Review.
-
Elucidation of the molecular basis of selective recognition uncovers the interaction site for the core domain of scorpion alpha-toxins on sodium channels.J Biol Chem. 2011 Oct 7;286(40):35209-17. doi: 10.1074/jbc.M111.259507. Epub 2011 Aug 8. J Biol Chem. 2011. PMID: 21832067 Free PMC article.
References
-
- Catterall W. A. From ionic currents to molecular mechanisms: the structure and function of voltage-gated sodium channels. Neuron. 2000;26:13–25. - PubMed
-
- Ulbricht W. Sodium channel inactivation: molecular determinants and modulation. Physiol. Rev. 2005;85:1271–1301. - PubMed
-
- Blumenthal K. M., Seibert A. L. Voltage-gated sodium channel toxins: poison, probes, and future promise. Cell. Biochem. Biophys. 2003;38:215–238. - PubMed
-
- Catterall W. A., Beress L. Sea anemone toxin and scorpion toxin share a common receptor site associated with the action potential sodium ionophore. J. Biol. Chem. 1978;253:7393–7396. - PubMed
-
- Little M. J., Zappia C., Gilles N., Connor M., Tyler M. I., Martin-Eauclaire M. F., Gordon D., Nicholson G. M. δ-Atracotoxins from Australian funnel-web spiders compete with scorpion α-toxin binding but differentially modulate alkaloid toxin activation of voltage-gated sodium channels. J. Biol. Chem. 1998;273:27076–27083. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources