

Dynamic circuitry for updating spatial representations. III. From neurons to behavior
- PMID: 17493922
- PMCID: PMC2383318
- DOI: 10.1152/jn.00330.2007
Dynamic circuitry for updating spatial representations. III. From neurons to behavior
Abstract
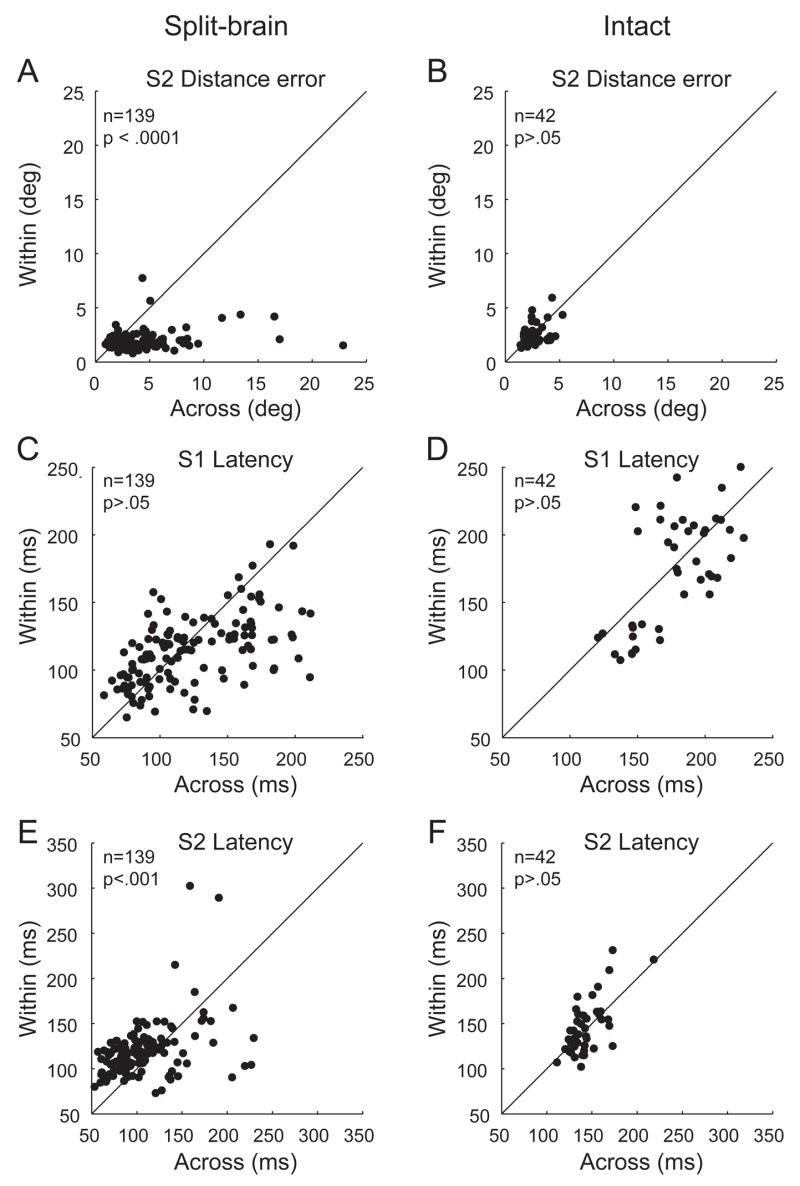
Each time the eyes move, the visual system must adjust internal representations to account for the accompanying shift in the retinal image. In the lateral intraparietal cortex (LIP), neurons update the spatial representations of salient stimuli when the eyes move. In previous experiments, we found that split-brain monkeys were impaired on double-step saccade sequences that required updating across visual hemifields, as compared to within hemifield. Here we describe a subsequent experiment to characterize the relationship between behavioral performance and neural activity in LIP in the split-brain monkey. We recorded from single LIP neurons while split-brain and intact monkeys performed two conditions of the double-step saccade task: one required across-hemifield updating and the other required within-hemifield updating. We found that, despite extensive experience with the task, the split-brain monkeys were significantly more accurate for within-hemifield than for across-hemifield sequences. In parallel, we found that population activity in LIP of the split-brain monkeys was significantly stronger for the within-hemifield than for the across-hemifield condition of the double-step task. In contrast, in the normal monkey, both the average behavioral performance and population activity showed no bias toward the within-hemifield condition. Finally, we found that the difference between within-hemifield and across-hemifield performance in the split-brain monkeys was reflected at the level of single-neuron activity in LIP. These findings indicate that remapping activity in area LIP is present in the split-brain monkey for the double-step task and covaries with spatial behavior on within-hemifield compared to across-hemifield sequences.
Figures










References
-
- Andersen RA. Handbook of Physiology: The Nervous System. Bethesda, MD: Williams Wekens; 1987. Inferior parietal lobule function in spatial perception and visuomotor integration; pp. 483–518.
-
- Assad JA. Neural coding of behavioral relevance in parietal cortex. Curr Opin Neurobiol. 2003;13:194–197. - PubMed
-
- Baizer JS, Bender DB. Comparison of saccadic eye movements in humans and macaques to single-step and double-step target movements. Vision Res. 1989;29:485–495. - PubMed
-
- Baker JT, Harper TM, Snyder LH. Spatial memory following shifts of gaze. I. Saccades to memorized world-fixed and gaze-fixed targets. J Neurophysiol. 2003;89:2564–2576. - PubMed
-
- Barash S, Bracewell RM, Fogassi L, Gnadt JW, Andersen RA. Saccade-related activity in the lateral intraparietal area. I. Temporal properties; comparison with area 7a. J Neurophysiol. 1991;66:1095–1108. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources