

TRPC3 channels are necessary for brain-derived neurotrophic factor to activate a nonselective cationic current and to induce dendritic spine formation
- PMID: 17494704
- PMCID: PMC2806846
- DOI: 10.1523/JNEUROSCI.5499-06.2007
TRPC3 channels are necessary for brain-derived neurotrophic factor to activate a nonselective cationic current and to induce dendritic spine formation
Abstract
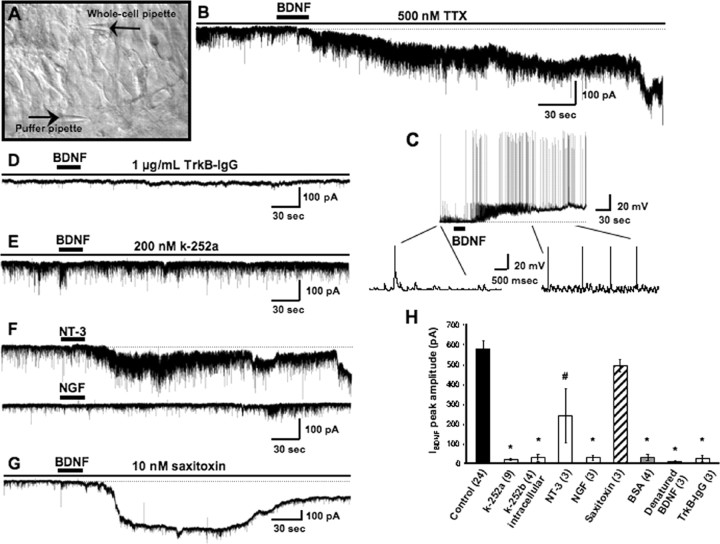
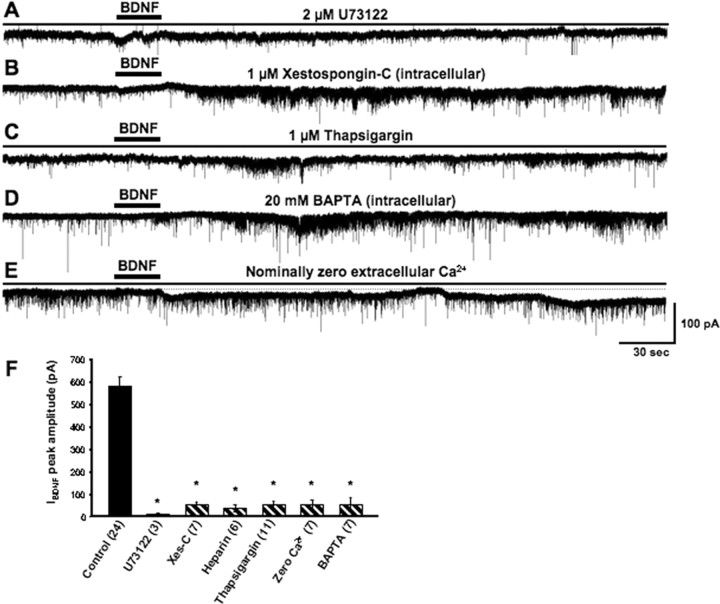
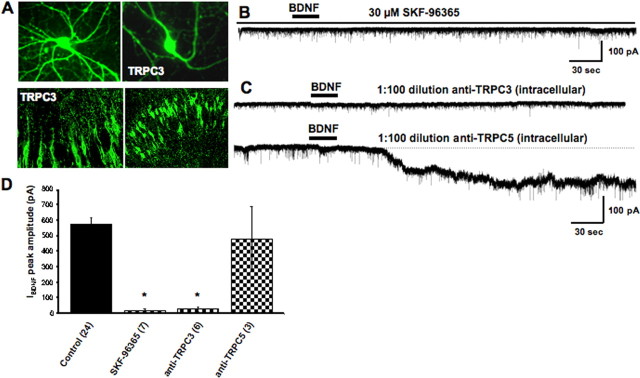
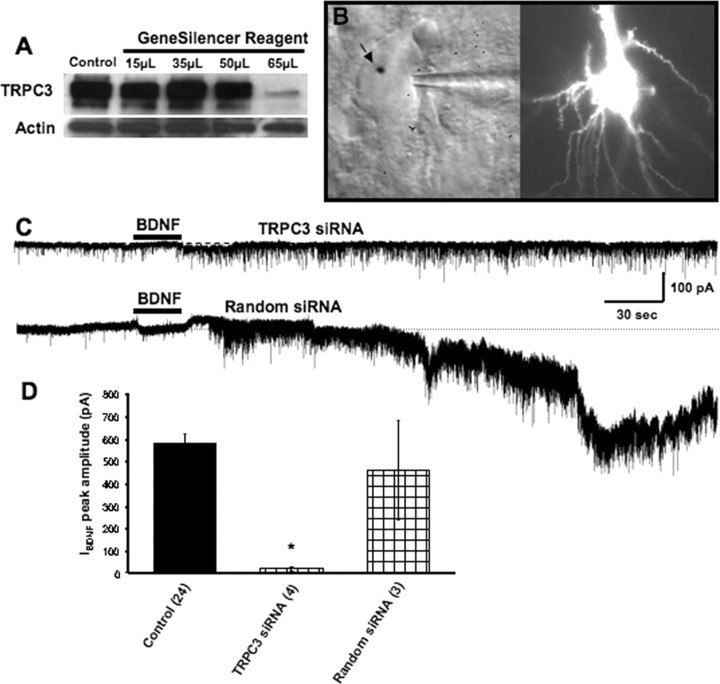
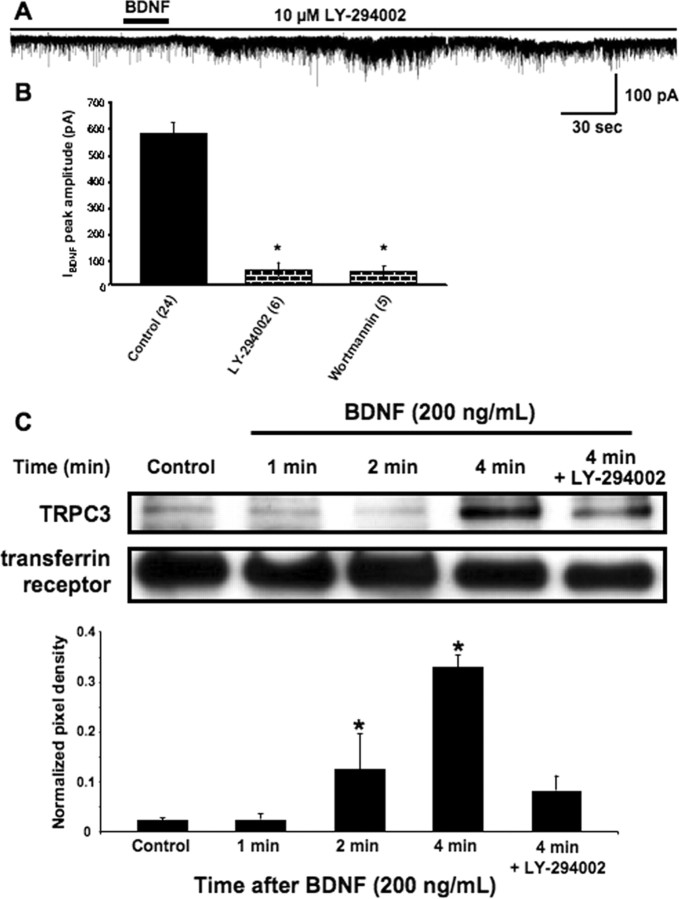
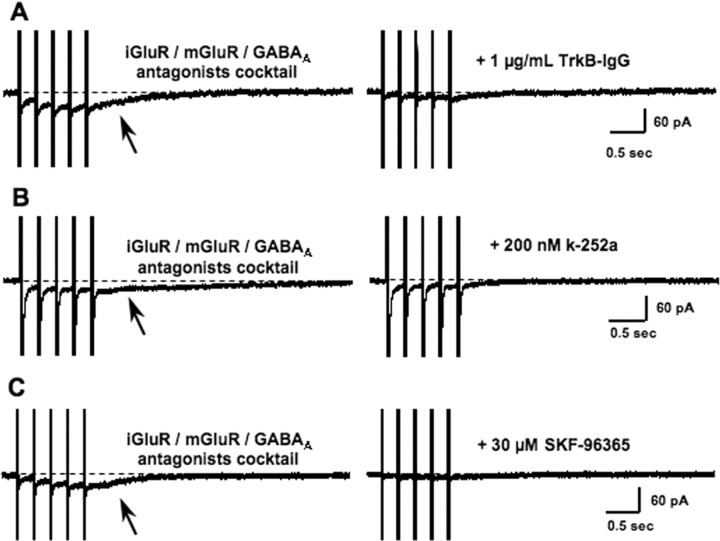
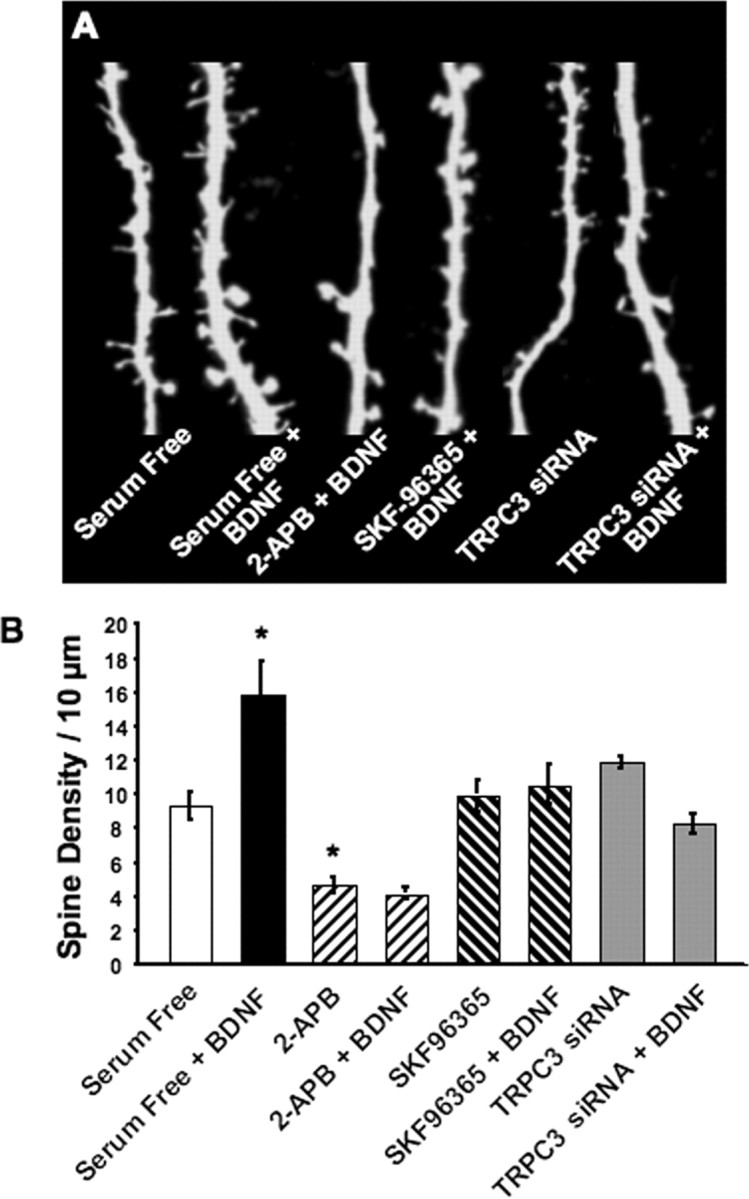
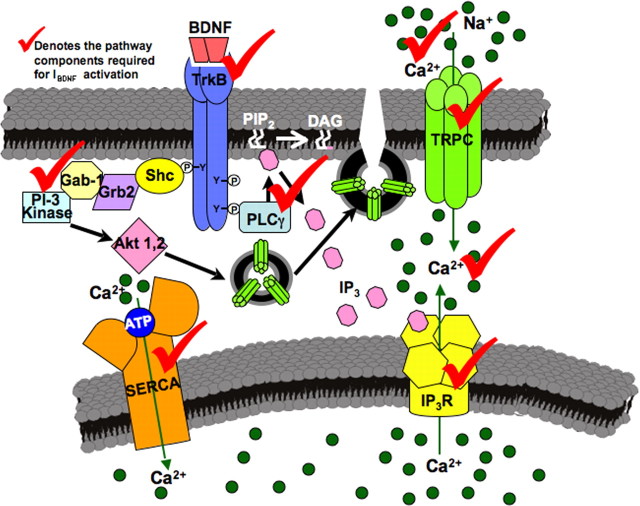
Brain-derived neurotrophic factor (BDNF) exerts prominent effects on hippocampal neurons, but the mechanisms that initiate its actions are poorly understood. We report here that BDNF evokes a slowly developing and sustained nonselective cationic current (I(BDNF)) in CA1 pyramidal neurons. These responses require phospholipase C, IP3 receptors, Ca2+ stores, and Ca2+ influx, suggesting the involvement of transient receptor potential canonical subfamily (TRPC) channels. Indeed, I(BDNF) is absent after small interfering RNA-mediated TRPC3 knockdown. The sustained kinetics of I(BDNF) appears to depend on phosphatidylinositol 3-kinase-mediated TRPC3 membrane insertion, as shown by surface biotinylation assays. Slowly emerging membrane currents after theta burst stimulation are sensitive to the scavenger TrkB-IgG and TRPC inhibitors, suggesting I(BDNF) activation by evoked released of endogenous, native BDNF. Last, TRPC3 channels are necessary for BDNF to increase dendritic spine density. Thus, TRPC channels emerge as novel mediators of BDNF-mediated dendritic remodeling through the activation of a slowly developing and sustained membrane depolarization.
Figures
References
-
- Agell N, Bachs O, Rocamora N, Villalonga P. Modulation of the Ras/Raf/MEK/ERK pathway by Ca2+, and calmodulin. Cell Signal. 2002;14:649–654. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous