

On the origin and evolutionary diversification of beetle horns
- PMID: 17494751
- PMCID: PMC1876444
- DOI: 10.1073/pnas.0701209104
On the origin and evolutionary diversification of beetle horns
Abstract
Many scarab beetles produce rigid projections from the body called horns. The exaggerated sizes of these structures and the staggering diversity of their forms have impressed biologists for centuries. Recent comparative studies using DNA sequence-based phylogenies have begun to reconstruct the historical patterns of beetle horn evolution. At the same time, developmental genetic experiments have begun to elucidate how beetle horns grow and how horn growth is modulated in response to environmental variables, such as nutrition. We bring together these two perspectives to show that they converge on very similar conclusions regarding beetle evolution. Horns do not appear to be difficult structures to gain or lose, and they can diverge both dramatically and rapidly in form. Although much of this work is still preliminary, we use available information to propose a conceptual developmental model for the major trajectories of beetle horn evolution. We illustrate putative mechanisms underlying the evolutionary origin of horns and the evolution of horn location, shape, allometry, and dimorphism.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
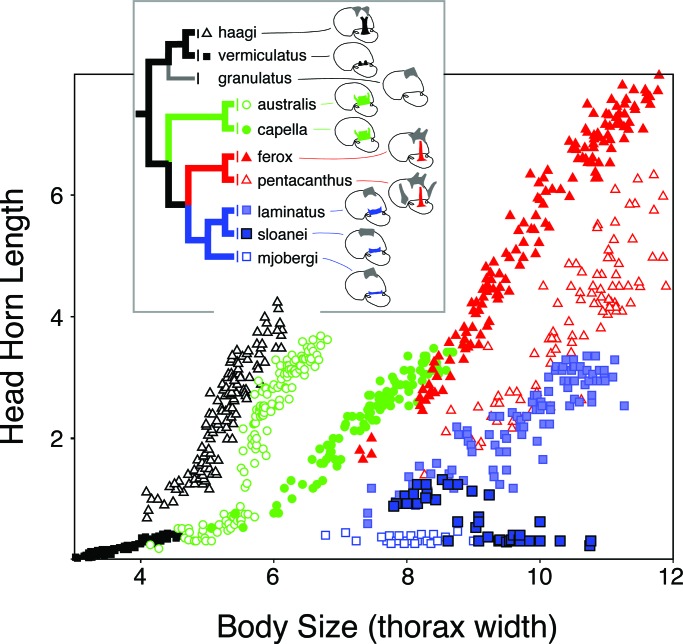
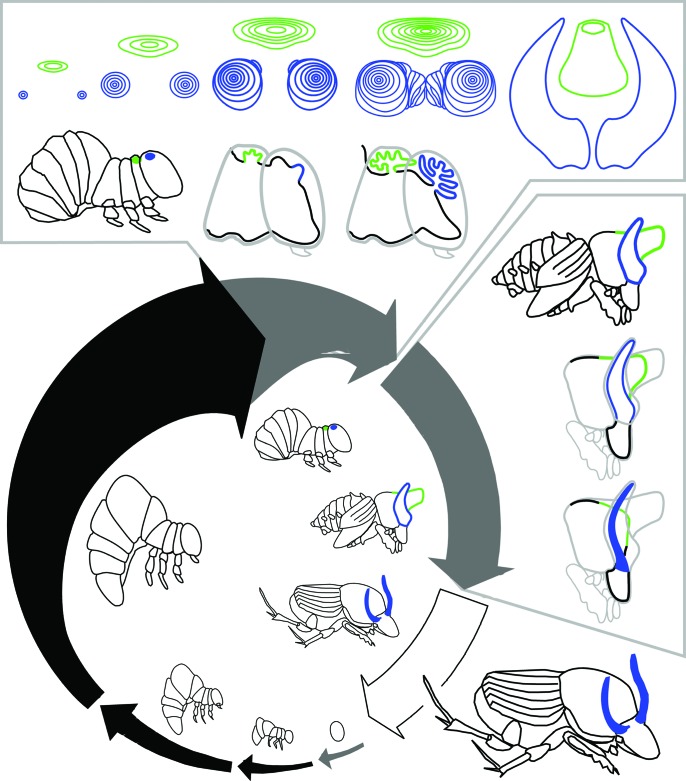
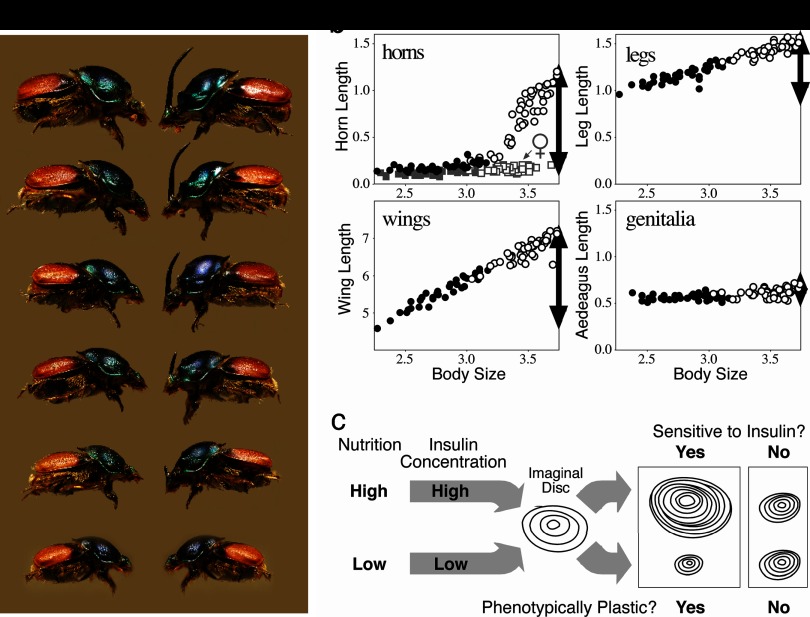
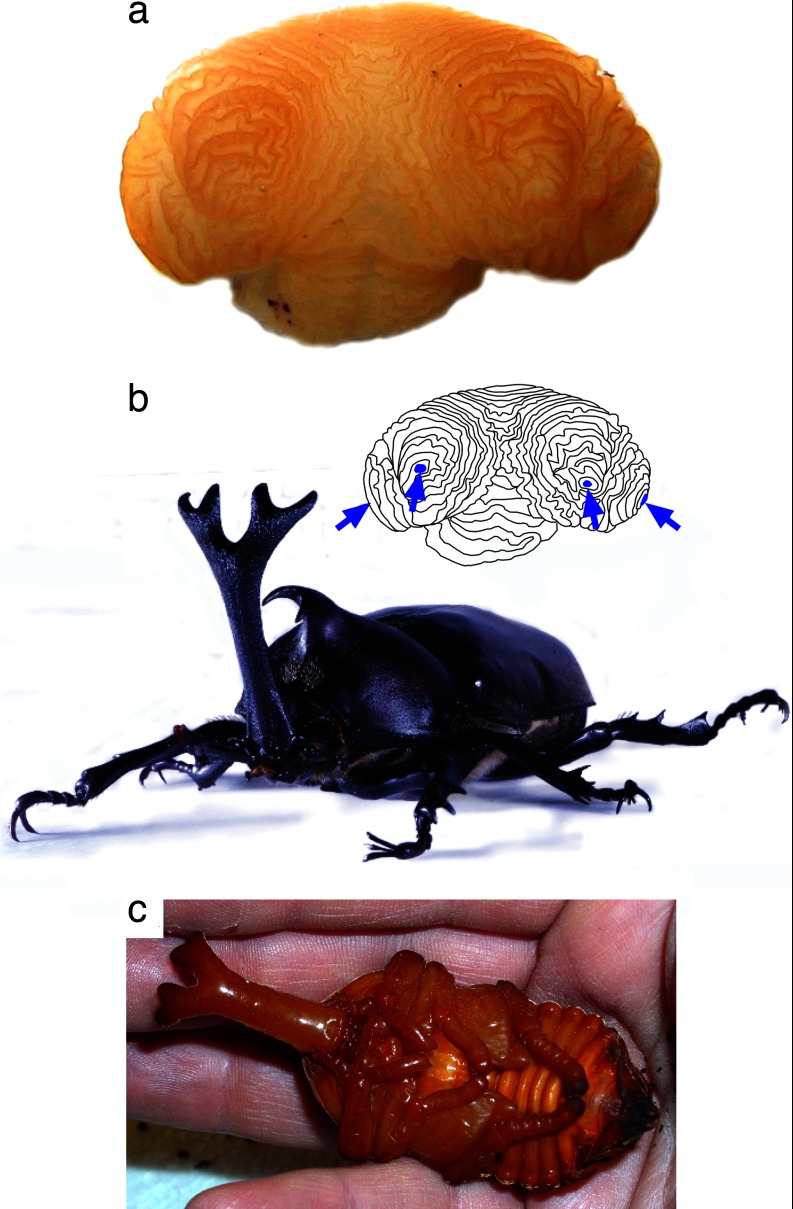
References
-
- Darwin C. On the Origin of Species by Means of Natural Selection. London: John Murray; 1859.
-
- Goldschmidt R. The Material Basis of Evolution. New Haven: Yale Univ Press; 1940.
-
- Mayr E. In: Evolution After Darwin. Tax S, editor. Chicago: Univ of Chicago Press; 1960. pp. 349–380.
-
- Müller GB, Wagner GP. Ann Rev Ecol Syst. 1991;22:229–256.
-
- Nitecki MH. Evolutionary Innovations. Chicago: Univ Chicago Press; 1990.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
