

Integrated network analysis identifies nitric oxide response networks and dihydroxyacid dehydratase as a crucial target in Escherichia coli
- PMID: 17494765
- PMCID: PMC1895976
- DOI: 10.1073/pnas.0610888104
Integrated network analysis identifies nitric oxide response networks and dihydroxyacid dehydratase as a crucial target in Escherichia coli
Abstract
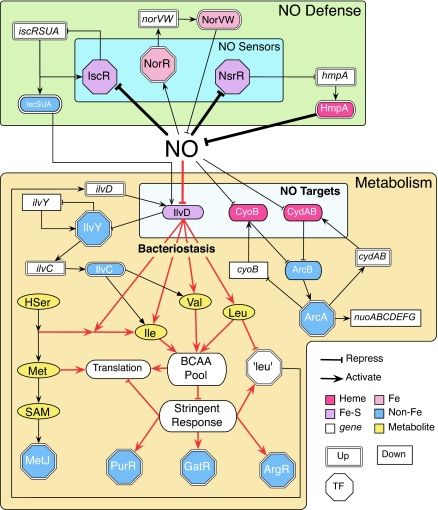
Nitric oxide (NO) is used by mammalian immune systems to counter microbial invasions and is produced by bacteria during denitrification. As a defense, microorganisms possess a complex network to cope with NO. Here we report a combined transcriptomic, chemical, and phenotypic approach to identify direct NO targets and construct the biochemical response network. In particular, network component analysis was used to identify transcription factors that are perturbed by NO. Such information was screened with potential NO reaction mechanisms and phenotypic data from genetic knockouts to identify active chemistry and direct NO targets in Escherichia coli. This approach identified the comprehensive E. coli NO response network and evinced that NO halts bacterial growth via inhibition of the branched-chain amino acid biosynthesis enzyme dihydroxyacid dehydratase. Because mammals do not synthesize branched-chain amino acids, inhibition of dihydroxyacid dehydratase may have served to foster the role of NO in the immune arsenal.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
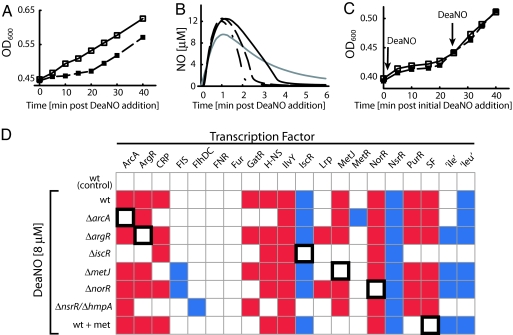
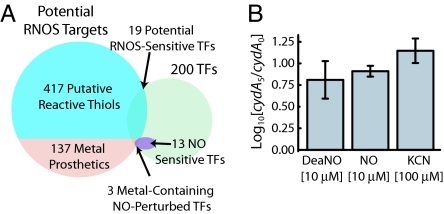
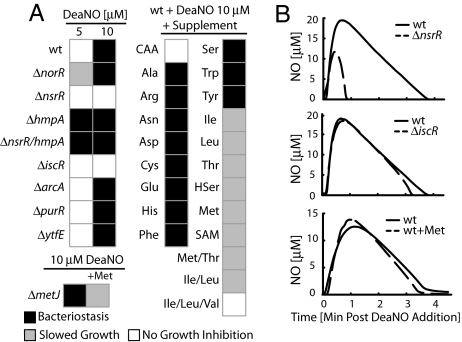
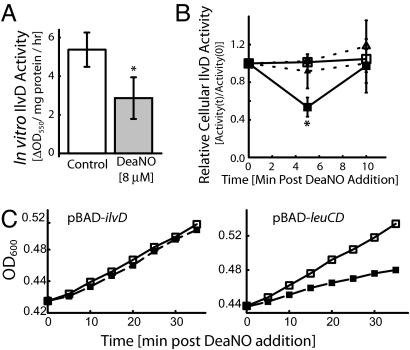
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
