

Extreme accumulation of nucleotides in simulated hydrothermal pore systems
- PMID: 17494767
- PMCID: PMC1890497
- DOI: 10.1073/pnas.0609592104
Extreme accumulation of nucleotides in simulated hydrothermal pore systems
Abstract
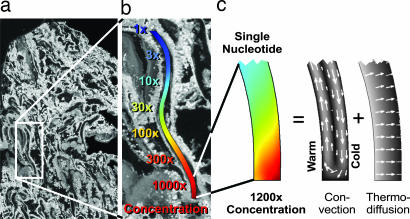
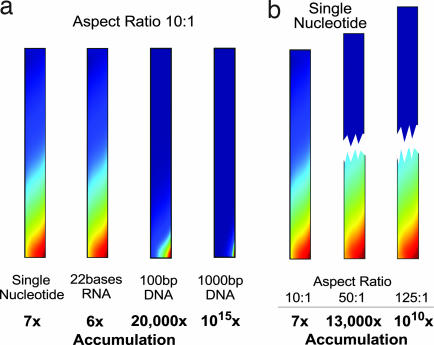
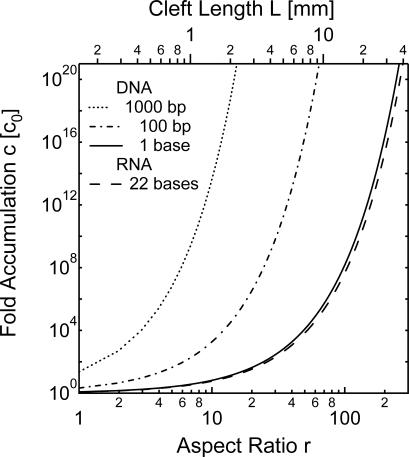
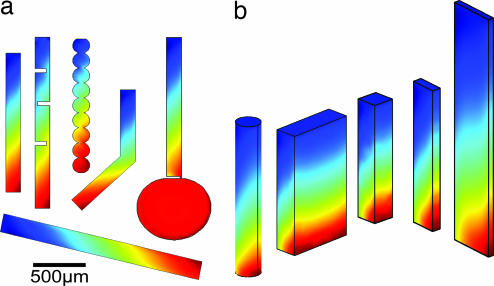
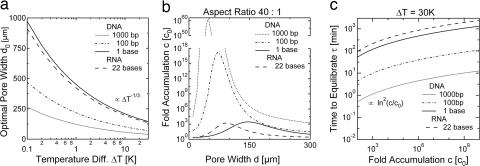
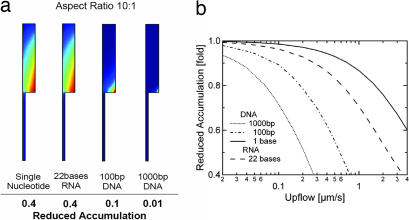
We simulate molecular transport in elongated hydrothermal pore systems influenced by a thermal gradient. We find extreme accumulation of molecules in a wide variety of plugged pores. The mechanism is able to provide highly concentrated single nucleotides, suitable for operations of an RNA world at the origin of life. It is driven solely by the thermal gradient across a pore. On the one hand, the fluid is shuttled by thermal convection along the pore, whereas on the other hand, the molecules drift across the pore, driven by thermodiffusion. As a result, millimeter-sized pores accumulate even single nucleotides more than 10(8)-fold into micrometer-sized regions. The enhanced concentration of molecules is found in the bulk water near the closed bottom end of the pore. Because the accumulation depends exponentially on the pore length and temperature difference, it is considerably robust with respect to changes in the cleft geometry and the molecular dimensions. Whereas thin pores can concentrate only long polynucleotides, thicker pores accumulate short and long polynucleotides equally well and allow various molecular compositions. This setting also provides a temperature oscillation, shown previously to exponentially replicate DNA in the protein-assisted PCR. Our results indicate that, for life to evolve, complicated active membrane transport is not required for the initial steps. We find that interlinked mineral pores in a thermal gradient provide a compelling high-concentration starting point for the molecular evolution of life.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
An RNA-making reactor for the origin of life.Proc Natl Acad Sci U S A. 2007 May 29;104(22):9105-6. doi: 10.1073/pnas.0702699104. Epub 2007 May 22. Proc Natl Acad Sci U S A. 2007. PMID: 17519331 Free PMC article. No abstract available.
References
-
- Miller SL. Science. 1953;15:528–529. - PubMed
-
- Drobner E, Huber H, Wächtershäuser G, Rose D, Stetter KO. Nature. 1990;346:742–744.
-
- Zubay G. Origins of Life on the Earth and in the Cosmos. 2nd Ed. New York: Academic; 2000.
-
- Ferris JP. Origins Life Evol Biosph. 2002;32:311–332. - PubMed
-
- Eigen M. Naturwissenschaften. 1971;58:465–523. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
