

Nudel modulates kinetochore association and function of cytoplasmic dynein in M phase
- PMID: 17494871
- PMCID: PMC1924840
- DOI: 10.1091/mbc.e06-04-0345
Nudel modulates kinetochore association and function of cytoplasmic dynein in M phase
Abstract
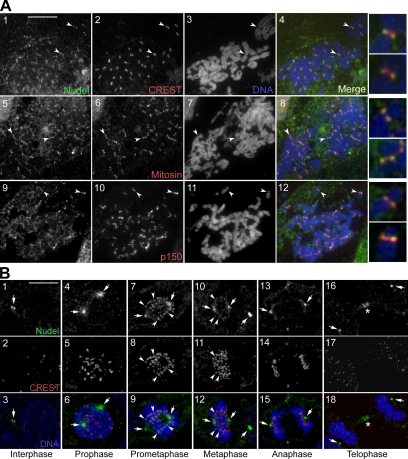
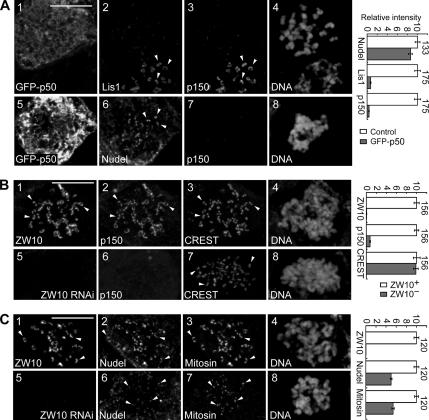
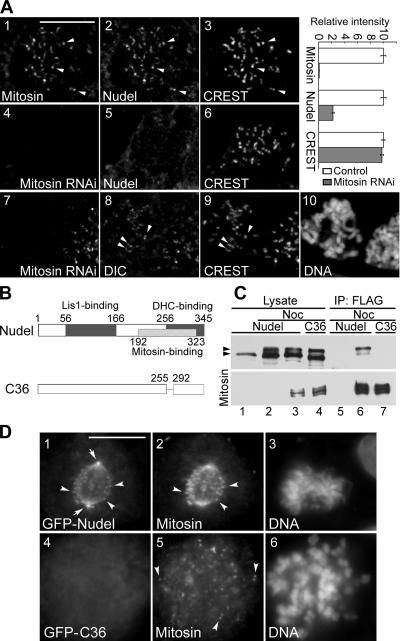
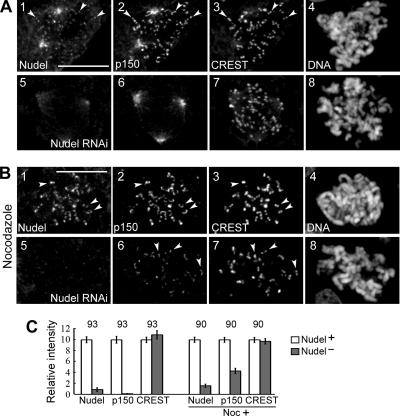
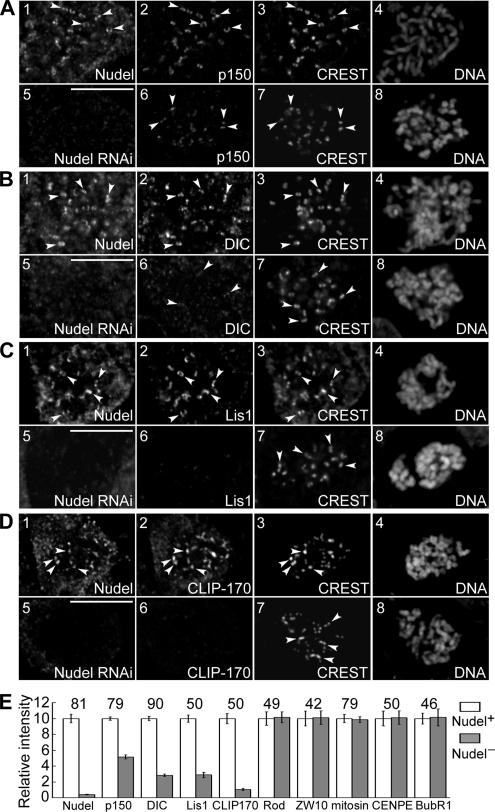
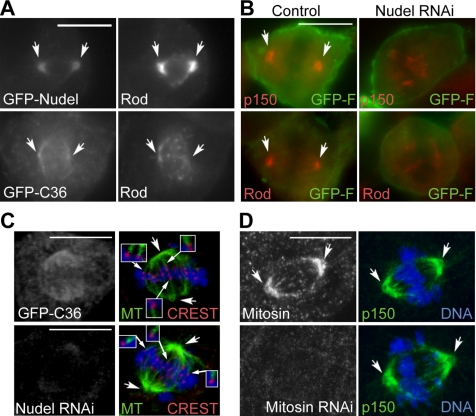
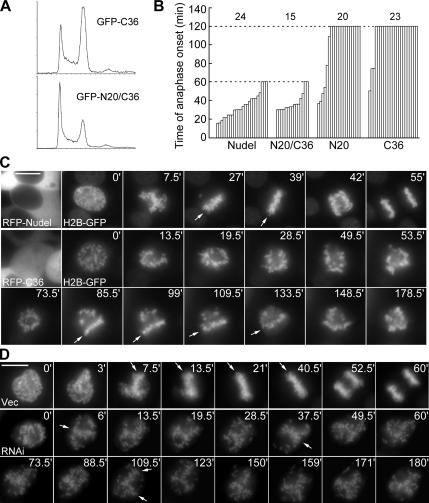
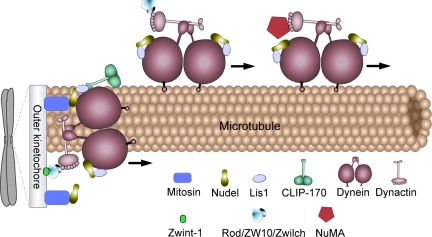
The microtubule-based motor cytoplasmic dynein/dynactin is a force generator at the kinetochore. It also transports proteins away from kinetochores to spindle poles. Regulation of such diverse functions, however, is poorly understood. We have previously shown that Nudel is critical for dynein-mediated protein transport, whereas mitosin, a kinetochore protein that binds Nudel, is involved in retention of kinetochore dynein/dynactin against microtubule-dependent stripping. Here we demonstrate that Nudel is required for robust localization of dynein/dynactin at the kinetochore. It localizes to kinetochores after nuclear envelope breakdown, depending mostly ( approximately 78%) on mitosin and slightly on dynein/dynactin. Depletion of Nudel by RNA interference (RNAi) or overexpression of its mutant incapable of binding either Lis1 or dynein heavy chain abolishes the kinetochore protein transport and mitotic progression. Similar to mitosin RNAi, Nudel RNAi also leads to increased stripping of kinetochore dynein/dynactin in the presence of microtubules. Taking together, our results suggest a dual role of kinetochore Nudel: it activates dynein-mediated protein transport and, when interacting with both mitosin and dynein, stabilizes kinetochore dynein/dynactin against microtubule-dependent stripping to facilitate the force generation function of the motor.
Figures
References
-
- Chan G. K., Jablonski S. A., Starr D. A., Goldberg M. L., Yen T. J. Human Zw10 and ROD are mitotic checkpoint proteins that bind to kinetochores. Nat. Cell Biol. 2000;2:944–947. - PubMed
-
- Cleveland D. W., Mao Y., Sullivan K. F. Centromeres and kinetochores: from epigenetics to mitotic checkpoint signaling. Cell. 2003;112:407–421. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
