

Changes in firing pattern of lateral geniculate neurons caused by membrane potential dependent modulation of retinal input through NMDA receptors
- PMID: 17495043
- PMCID: PMC2075279
- DOI: 10.1113/jphysiol.2007.131540
Changes in firing pattern of lateral geniculate neurons caused by membrane potential dependent modulation of retinal input through NMDA receptors
Abstract
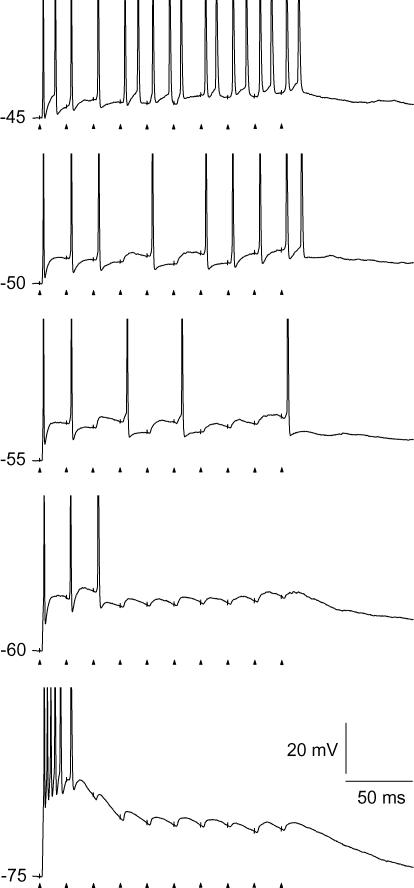
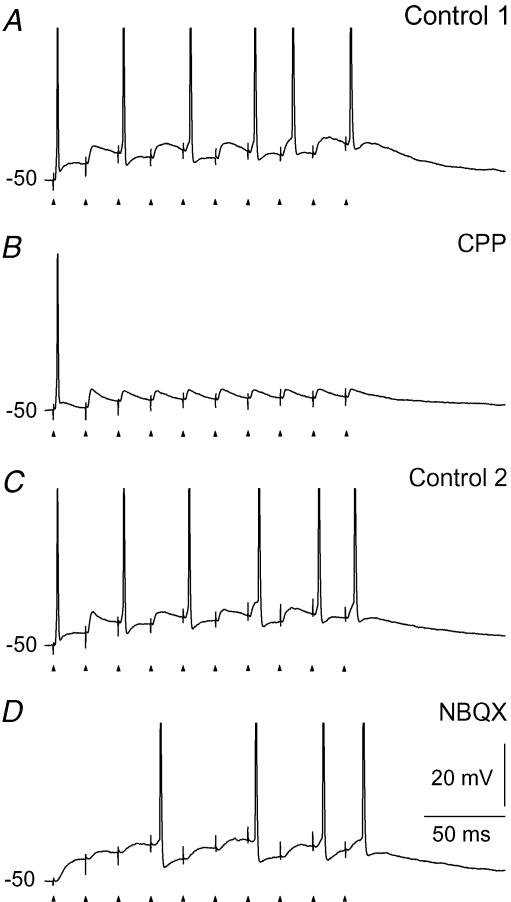
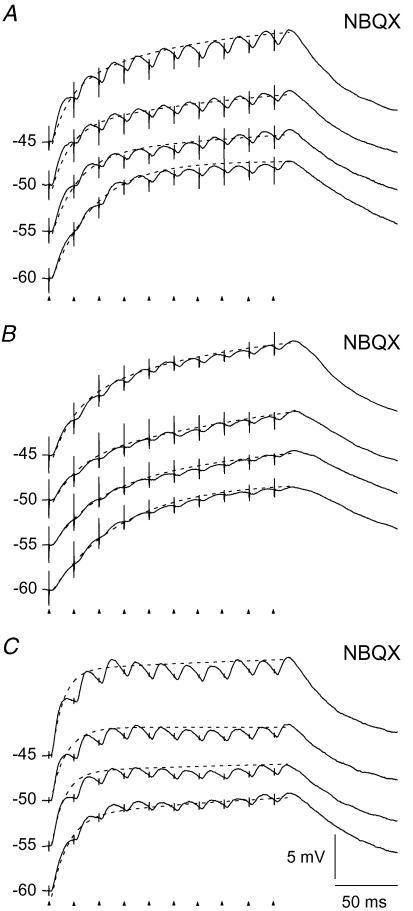
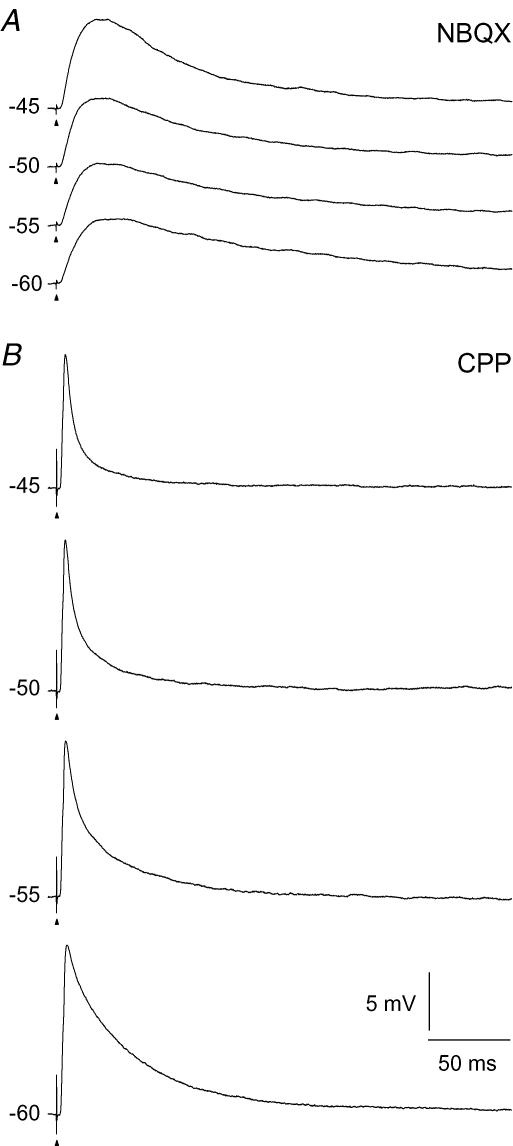
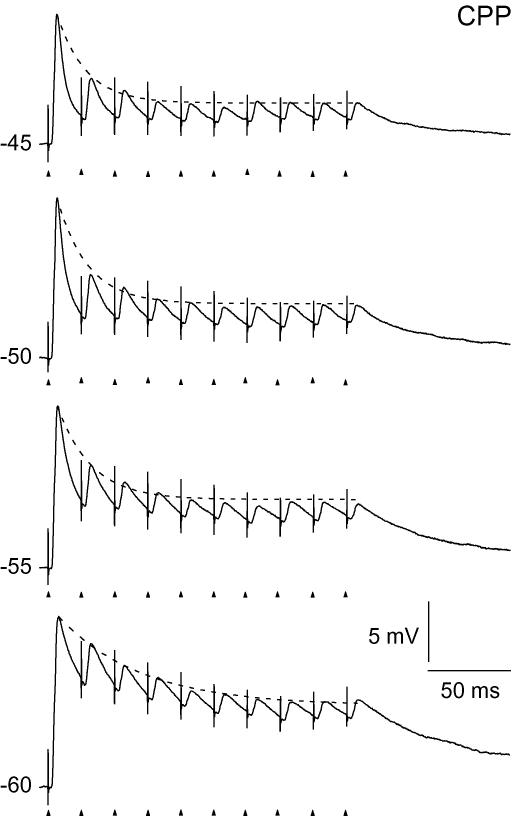
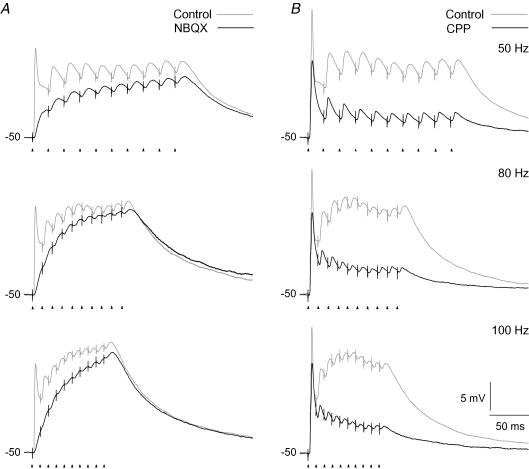
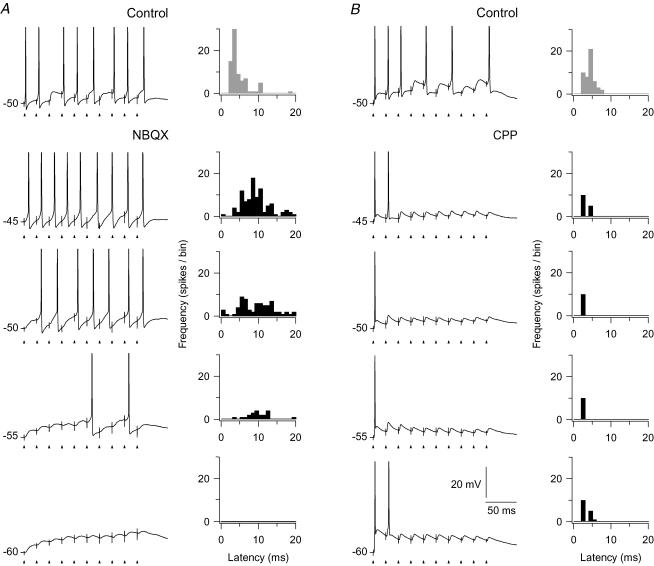
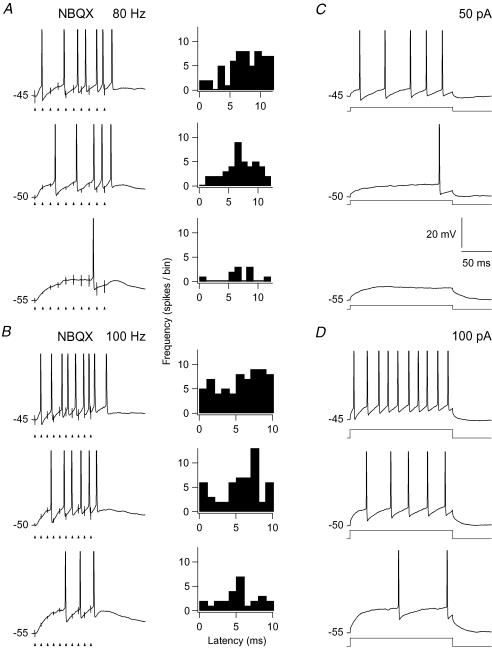
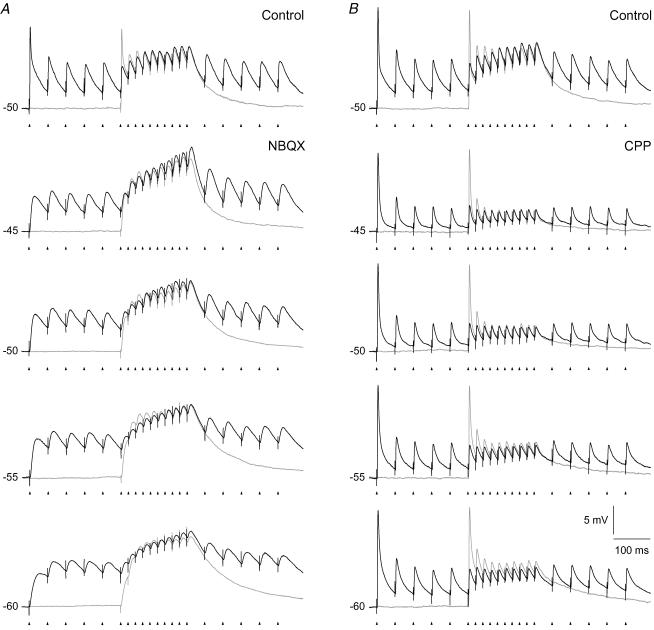
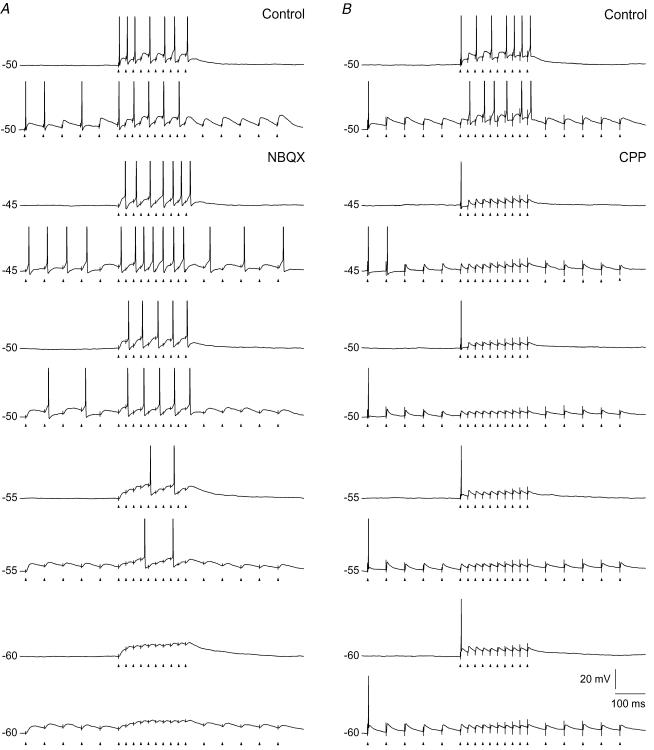
An optimal visual stimulus flashed on the receptive field of a retinal ganglion cell typically evokes a strong transient response followed by weaker sustained firing. Thalamocortical (TC) neurons in the dorsal lateral geniculate nucleus, which receive their sensory input from retina, respond similarly except that the gain, in particular of the sustained component, changes with level of arousal. Several lines of evidence suggest that retinal input to TC neurons through NMDA receptors plays a key role in generation of the sustained response, but the mechanisms for the state-dependent variation in this component are unclear. We used a slice preparation to study responses of TC neurons evoked by trains of electrical pulses to the retinal afferents at frequencies in the range of visual responses in vivo. Despite synaptic depression, the pharmacologically isolated NMDA component gave a pronounced build-up of depolarization through temporal summation of the NMDA receptor mediated EPSPs. This depolarization could provide sustained firing, the frequency of which depended on the holding potential. We suggest that the variation of sustained response in vivo is caused mainly by the state-dependent modulation of the membrane potential of TC neurons which shifts the NMDA receptor mediated depolarization closer to or further away from the firing threshold. The pharmacologically isolated AMPA receptor EPSPs were rather ineffective in spike generation. However, together with the depolarization evoked by the NMDA component, the AMPA component contributed significantly to spike generation, and was necessary for the precise timing of the generated spikes.
Figures

References
-
- Balkema GW, Pinto LH. Electrophysiology of retinal ganglion cells in the mouse: a study of a normally pigmented mouse and a congenic hypopigmentation mutant, pearl. J Neurophysiol. 1982;48:968–980. - PubMed
-
- Blitz DM, Regehr WG. Retinogeniculate synaptic properties controlling spike number and timing in relay neurons. J Neurophysiol. 2003;90:2438–2450. - PubMed
-
- Blitz DM, Regehr WG. Timing and specificity of feed-forward inhibition within the LGN. Neuron. 2005;45:917–928. - PubMed
-
- Chen CF, Blitz DM, Regehr WG. Contributions of receptor desensitization and saturation to plasticity at the retinogeniculate synapse. Neuron. 2002;33:779–788. - PubMed
-
- Coenen AML, Vendrik AJH. Determination of the transfer ratio of cat's geniculate neurons through quasi-intracellular recordings and the relation with the level of alertness. Exp Brain Res. 1972;14:227–242. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources