

The role of leaf lobation in elongation responses to shade in the rosette-forming forb Serratula tinctoria (Asteraceae)
- PMID: 17495981
- PMCID: PMC2735293
- DOI: 10.1093/aob/mcm074
The role of leaf lobation in elongation responses to shade in the rosette-forming forb Serratula tinctoria (Asteraceae)
Abstract
Background and aims: Lobed leaves are considered selectively advantageous in conditions of high irradiance. However, most studies have involved woody species, with only a few considering the role of leaf lobation in herbaceous rosette species. In this study, it is hypothesized that, in addition to its adaptive value in high light, leaf lobation may add to the function of petioles as vertical spacers in herbaceous species in conditions of strong competition for light.
Methods: To test this hypothesis, leaf development was examined under seasonally changing natural light conditions and a field experiment was conducted in which light climate was manipulated in a wooded meadow population of Serratula tinctoria.
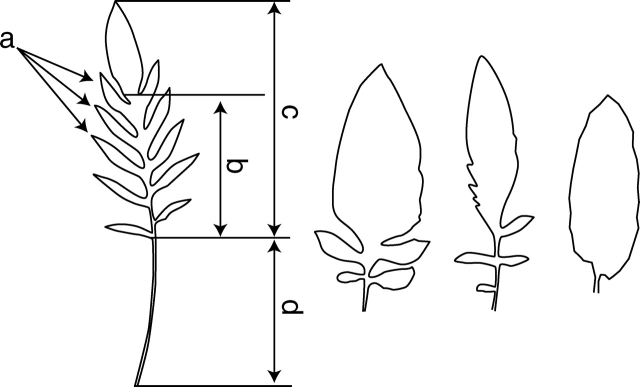
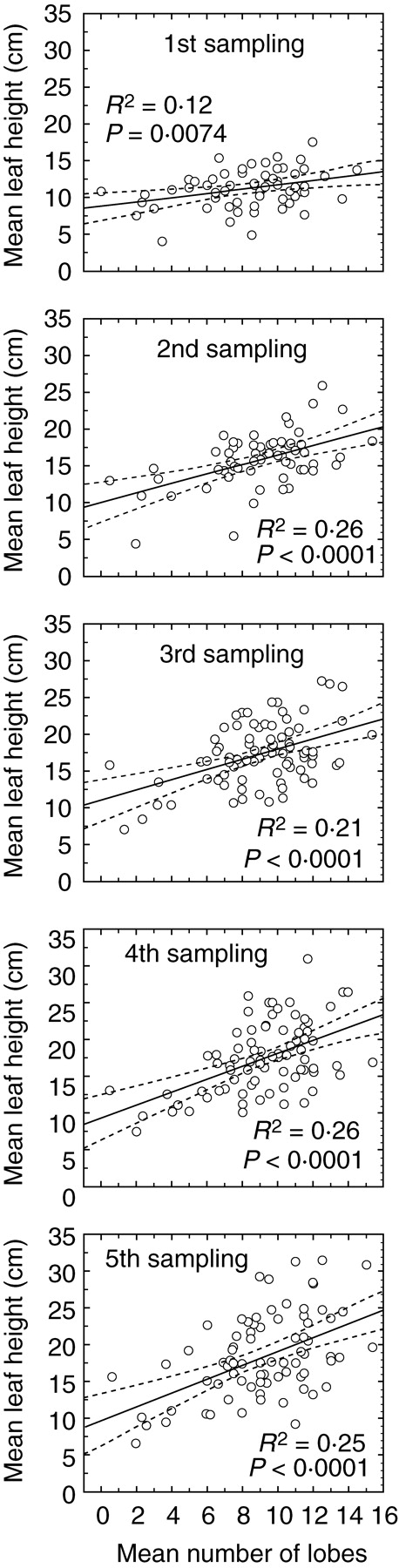
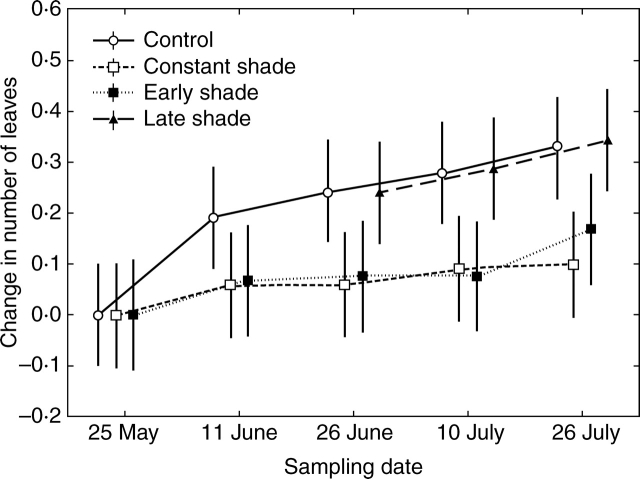
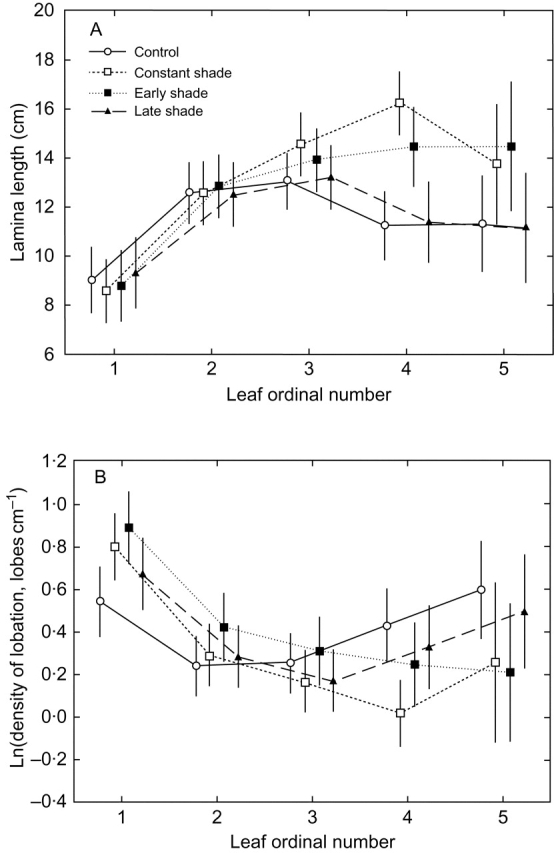
Key results: No changes in leaf lobation were observed in response to experimental shading or different natural light conditions. However, in tall herbaceous vegetation, plants with highly lobed leaves achieved significantly greater vertical elongation than plants with less-lobed leaves. In contrast to herbaceous shade, tree shade had no effect on leaf elongation, suggesting differential responsiveness to competition from neighbouring herbs versus overhead shade. In shading treatments, imposed shade could only be responded to by the elongation of leaves that were produced late in development.
Conclusions: The results show that extensive leaf lobation can enable greater leaf elongation in response to shade from surrounding herbaceous vegetation. The different morphological responses displayed by Serratula tinctoria to different types of shade demonstrate the importance of critically assessing experimental designs when investigating phenotypic plasticity in response to shade.
Figures


Similar articles
-
A dynamic analysis of the shade-induced plasticity in Arabidopsis thaliana rosette leaf development reveals new components of the shade-adaptative response.Ann Bot. 2006 Mar;97(3):443-52. doi: 10.1093/aob/mcj047. Epub 2005 Dec 21. Ann Bot. 2006. PMID: 16371443 Free PMC article.
-
Differential influence of clonal integration on morphological and growth responses to light in two invasive herbs.PLoS One. 2012;7(4):e35873. doi: 10.1371/journal.pone.0035873. Epub 2012 Apr 27. PLoS One. 2012. PMID: 22558248 Free PMC article.
-
Plasticity in seedling morphology, biomass allocation and physiology among ten temperate tree species in response to shade is related to shade tolerance and not leaf habit.Plant Biol (Stuttg). 2017 Mar;19(2):172-182. doi: 10.1111/plb.12531. Epub 2017 Jan 5. Plant Biol (Stuttg). 2017. PMID: 27981788
-
Photomorphogenesis of leaves: shade-avoidance and differentiation of sun and shade leaves.Photochem Photobiol Sci. 2005 Sep;4(9):770-4. doi: 10.1039/b418440h. Epub 2005 May 4. Photochem Photobiol Sci. 2005. PMID: 16121290 Review.
-
Plant Responses to Vegetation Proximity: A Whole Life Avoiding Shade.Front Plant Sci. 2016 Feb 29;7:236. doi: 10.3389/fpls.2016.00236. eCollection 2016. Front Plant Sci. 2016. PMID: 26973679 Free PMC article. Review.
Cited by
-
Overexpression of the Liriodendron tulipifera BOP2 Gene (LtuBOP2) Affects Leaf Margin Development in Transgenic Arabidopsis thaliana.Int J Mol Sci. 2023 Feb 7;24(4):3262. doi: 10.3390/ijms24043262. Int J Mol Sci. 2023. PMID: 36834696 Free PMC article.
-
Belowground plant competition: uncoupling root response strategies of peas.Proc Biol Sci. 2024 Jul;291(2027):20240673. doi: 10.1098/rspb.2024.0673. Epub 2024 Jul 31. Proc Biol Sci. 2024. PMID: 39079667 Free PMC article.
-
Homeodomain leucine zipper protein controls the lobed leaf formation by modulating auxin distribution in watermelon.Theor Appl Genet. 2025 Jun 20;138(7):156. doi: 10.1007/s00122-025-04931-x. Theor Appl Genet. 2025. PMID: 40540203
-
Ectopic Expression of BcCUC2 Involved in Sculpting the Leaf Margin Serration in Arabidopsis thaliana.Genes (Basel). 2023 Jun 15;14(6):1272. doi: 10.3390/genes14061272. Genes (Basel). 2023. PMID: 37372452 Free PMC article.
-
Decision-making in plants under competition.Nat Commun. 2017 Dec 21;8(1):2235. doi: 10.1038/s41467-017-02147-2. Nat Commun. 2017. PMID: 29269832 Free PMC article.
References
-
- Collins RP, Jones MB. The effects of temperature on leaf growth in Cyperus longus, a temperate C4 species. Annals of Botany. 1988;61:355–362.
-
- Corré WJ. Growth and morphogenesis of sun and shade plants. II. The influence of light quality. Acta Botanica Neerlandica. 1983;32:185–202.
-
- DeWitt TJ, Sih A, Wilson DS. Costs and limits of phenotypic plasticity. Trends in Ecology and Evolution. 1998;13:77–81. - PubMed
-
- Dudley SA, Schmitt J. Genetic differentiation in morphological responses to simulated foliage shade between populations of Impatiens capensis from open and woodland sites. Functional Ecology. 1995;9:655–666.
-
- Givnish TJ. On the adaptive significance of compound leaves, with particular reference to tropical trees. In: Tomlinson PB, Zimmerman MH, editors. Tropical trees as living systems. Cambidge: Cambridge University Press; 1978. pp. 351–380.
