

Identical, independent, and opposing roles of ppGpp and DksA in Escherichia coli
- PMID: 17496080
- PMCID: PMC1951846
- DOI: 10.1128/JB.00330-07
Identical, independent, and opposing roles of ppGpp and DksA in Escherichia coli
Abstract
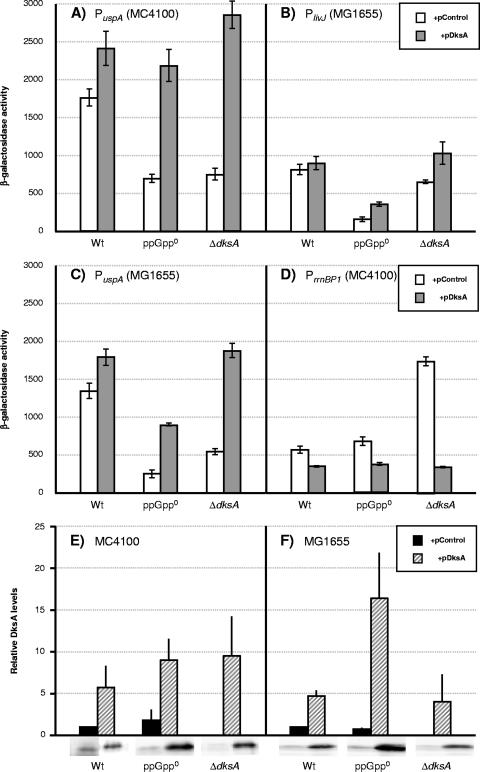
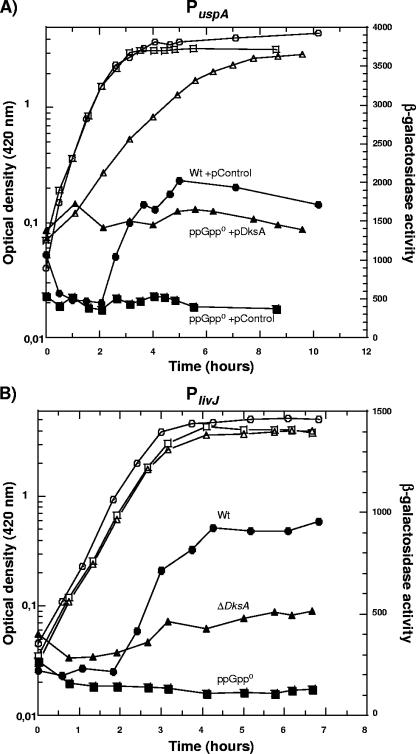
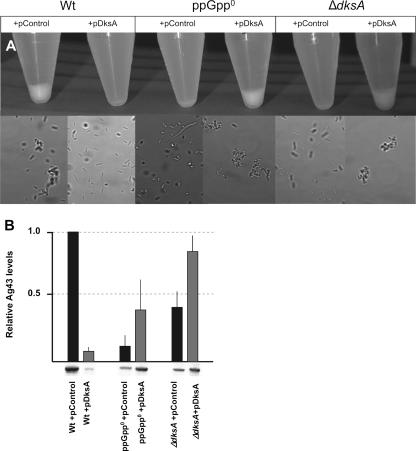
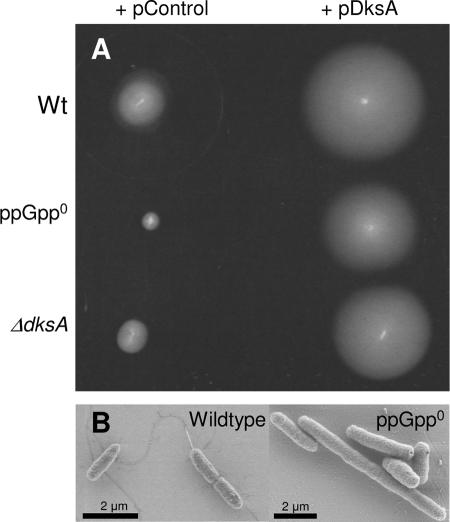
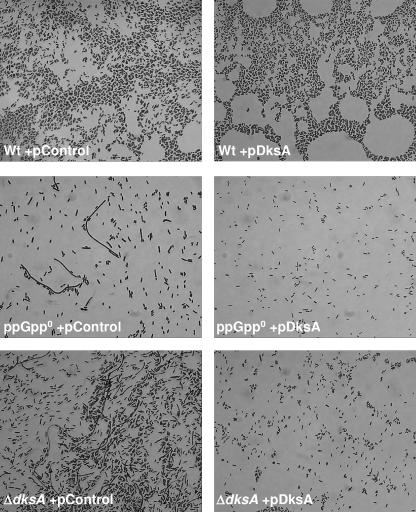
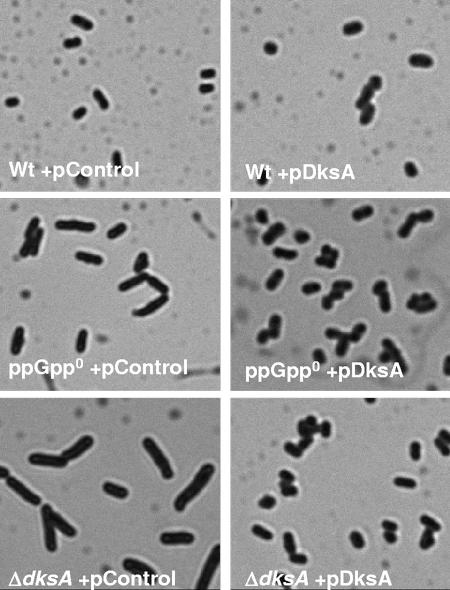
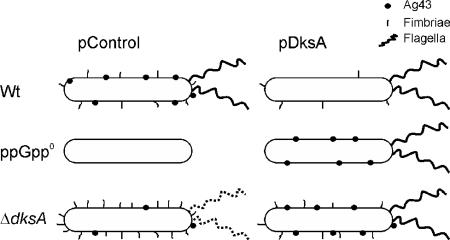
The recent discovery that the protein DksA acts as a coregulator of genes controlled by ppGpp led us to investigate the similarities and differences between the relaxed phenotype of a ppGpp-deficient mutant and the phenotype of a strain lacking DksA. We demonstrate that the absence of DksA and ppGpp has similar effects on many of the observed phenotypes but that DksA and ppGpp also have independent and sometimes opposing roles in the cell. Specifically, we show that overexpression of DksA can compensate for the loss of ppGpp with respect to transcription of the promoters P(uspA), P(livJ), and P(rrnBP1) as well as amino acid auxotrophy, cell-cell aggregation, motility, filamentation, and stationary phase morphology, suggesting that DksA can function without ppGpp in regulating gene expression. In addition, ppGpp and DksA have opposing effects on adhesion. In the course of our analysis, we also discovered new features of the relaxed mutant, namely, defects in cell-cell aggregation and motility.
Figures

References
-
- Aberg, A., V. Shingler, and C. Balsalobre. 2006. (p)ppGpp regulates type 1 fimbriation of Escherichia coli by modulating the expression of the site-specific recombinase FimB. Mol. Microbiol. 60:1520-1533. - PubMed
-
- Albertson, N. H., and T. Nystrom. 1994. Effects of starvation for exogenous carbon on functional mRNA stability and rate of peptide chain elongation in Escherichia coli. FEMS Microbiol. Lett. 117:181-187. - PubMed
-
- Artsimovitch, I., V. Patlan, S. Sekine, M. N. Vassylyeva, T. Hosaka, K. Ochi, S. Yokoyama, and D. G. Vassylyev. 2004. Structural basis for transcription regulation by alarmone ppGpp. Cell 117:299-310. - PubMed
-
- Barker, M. M., T. Gaal, C. A. Josaitis, and R. L. Gourse. 2001. Mechanism of regulation of transcription initiation by ppGpp. I. Effects of ppGpp on transcription initiation in vivo and in vitro. J. Mol. Biol. 305:673-688. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
