

Identification of a bipartite jasmonate-responsive promoter element in the Catharanthus roseus ORCA3 transcription factor gene that interacts specifically with AT-Hook DNA-binding proteins
- PMID: 17496112
- PMCID: PMC1914126
- DOI: 10.1104/pp.107.096115
Identification of a bipartite jasmonate-responsive promoter element in the Catharanthus roseus ORCA3 transcription factor gene that interacts specifically with AT-Hook DNA-binding proteins
Abstract
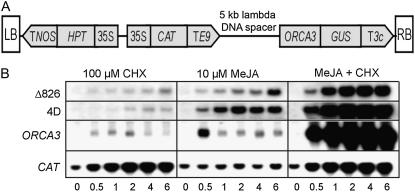
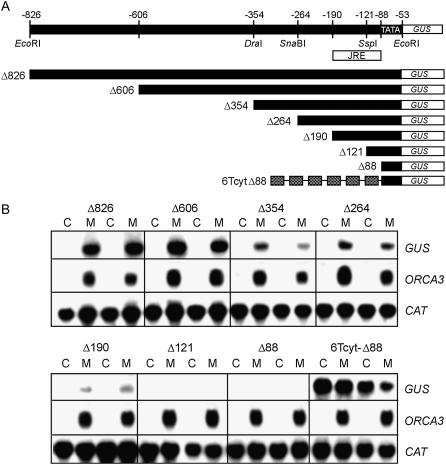
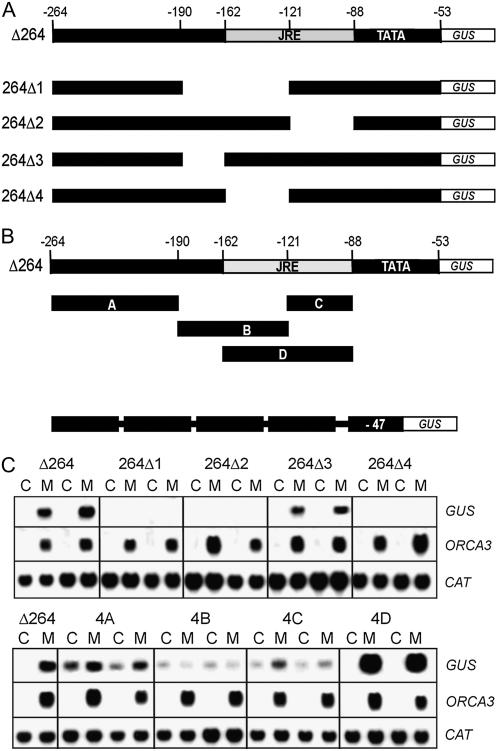
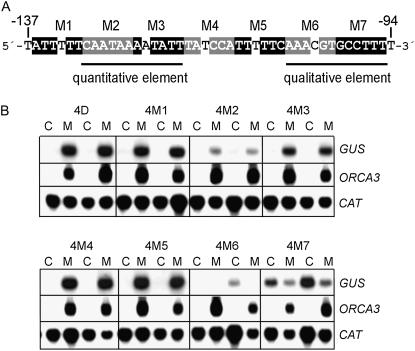
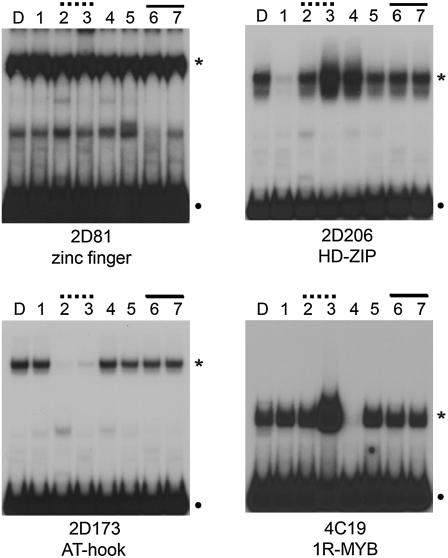
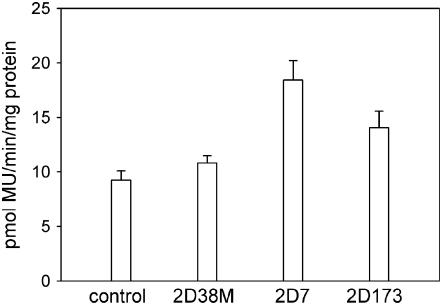
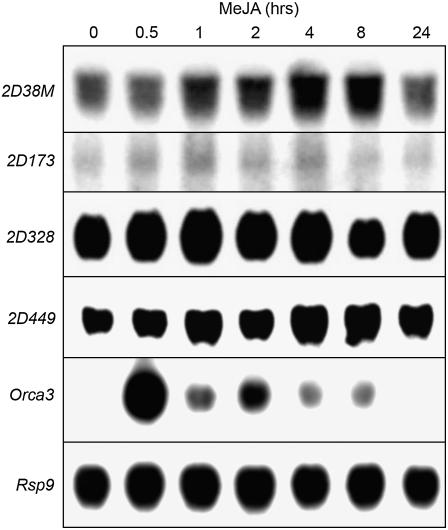
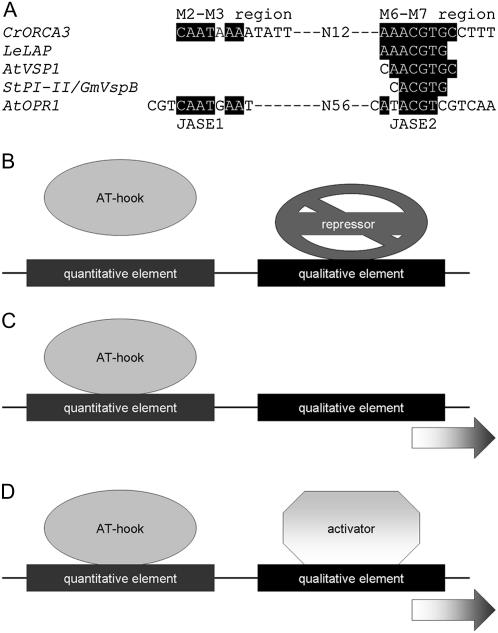
Jasmonates are plant signaling molecules that play key roles in defense against certain pathogens and insects, among others, by controlling the biosynthesis of protective secondary metabolites. In Catharanthus roseus, the APETALA2-domain transcription factor ORCA3 is involved in the jasmonate-responsive activation of terpenoid indole alkaloid biosynthetic genes. ORCA3 gene expression is itself induced by jasmonate. By loss- and gain-of-function experiments, we located a 74-bp region within the ORCA3 promoter, which contains an autonomous jasmonate-responsive element (JRE). The ORCA3 JRE is composed of two important sequences: a quantitative sequence responsible for a high level of expression and a qualitative sequence that appears to act as an on/off switch in response to methyl jasmonate. We isolated 12 different DNA-binding proteins having one of four different types of DNA-binding domains, using the ORCA3 JRE as bait in a yeast (Saccharomyces cerevisiae) one-hybrid transcription factor screening. The binding of one class of proteins bearing a single AT-hook DNA-binding motif was affected by mutations in the quantitative sequence within the JRE. Two of the AT-hook proteins tested had a weak activating effect on JRE-mediated reporter gene expression, suggesting that AT-hook family members may be involved in determining the level of expression of ORCA3 in response to jasmonate.
Figures
Similar articles
-
The basic helix-loop-helix transcription factor CrMYC2 controls the jasmonate-responsive expression of the ORCA genes that regulate alkaloid biosynthesis in Catharanthus roseus.Plant J. 2011 Jul;67(1):61-71. doi: 10.1111/j.1365-313X.2011.04575.x. Epub 2011 Apr 26. Plant J. 2011. PMID: 21401746
-
The jasmonate-responsive element from the ORCA3 promoter from Catharanthus roseus is active in Arabidopsis and is controlled by the transcription factor AtMYC2.Plant Cell Physiol. 2011 Mar;52(3):578-87. doi: 10.1093/pcp/pcr016. Epub 2011 Feb 8. Plant Cell Physiol. 2011. PMID: 21306988
-
The jasmonate-inducible AP2/ERF-domain transcription factor ORCA3 activates gene expression via interaction with a jasmonate-responsive promoter element.Plant J. 2001 Jan;25(1):43-53. doi: 10.1046/j.1365-313x.2001.00932.x. Plant J. 2001. PMID: 11169181
-
ORCAnization of jasmonate-responsive gene expression in alkaloid metabolism.Trends Plant Sci. 2001 May;6(5):212-9. doi: 10.1016/s1360-1385(01)01924-0. Trends Plant Sci. 2001. PMID: 11335174 Review.
-
Transcription factors: tools to engineer the production of pharmacologically active plant metabolites.Trends Pharmacol Sci. 2002 Dec;23(12):563-9. doi: 10.1016/s0165-6147(02)02098-9. Trends Pharmacol Sci. 2002. PMID: 12457774 Review.
Cited by
-
The Arabidopsis thaliana STYLISH1 protein acts as a transcriptional activator regulating auxin biosynthesis.Plant Cell. 2010 Feb;22(2):349-63. doi: 10.1105/tpc.108.064816. Epub 2010 Feb 12. Plant Cell. 2010. PMID: 20154152 Free PMC article.
-
Expression of Terpenoids 1, a glandular trichome-specific transcription factor from tomato that activates the terpene synthase 5 promoter.Plant Mol Biol. 2014 Feb;84(3):345-57. doi: 10.1007/s11103-013-0142-0. Epub 2013 Oct 20. Plant Mol Biol. 2014. PMID: 24142382
-
SUPPRESSOR OF PHYTOCHROME B-4 #3 reduces the expression of PIF-activated genes and increases expression of growth repressors to regulate hypocotyl elongation in short days.BMC Plant Biol. 2022 Aug 15;22(1):399. doi: 10.1186/s12870-022-03737-z. BMC Plant Biol. 2022. PMID: 35965321 Free PMC article.
-
Abscisic acid and other plant hormones: Methods to visualize distribution and signaling.Bioessays. 2015 Dec;37(12):1338-49. doi: 10.1002/bies.201500115. Bioessays. 2015. PMID: 26577078 Free PMC article. Review.
-
Identification of JAZ-interacting MYC transcription factors involved in latex drainage in Hevea brasiliensis.Sci Rep. 2018 Jan 17;8(1):909. doi: 10.1038/s41598-018-19206-3. Sci Rep. 2018. PMID: 29343866 Free PMC article.
References
-
- Chatel G, Montiel G, Pré M, Memelink J, Thiersault M, Saint-Pierre B, Doireau P, Gantet P (2003) CrMYC, a Catharanthus roseus elicitor- and jasmonate-responsive bHLH transcription factor that binds the G-box element of the strictosidine synthase gene promoter. J Exp Bot 54 2587–2588 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
