

Biochemical and crystallographic analysis of substrate binding and conformational changes in acetyl-CoA synthetase
- PMID: 17497934
- PMCID: PMC2536627
- DOI: 10.1021/bi6026506
Biochemical and crystallographic analysis of substrate binding and conformational changes in acetyl-CoA synthetase
Abstract
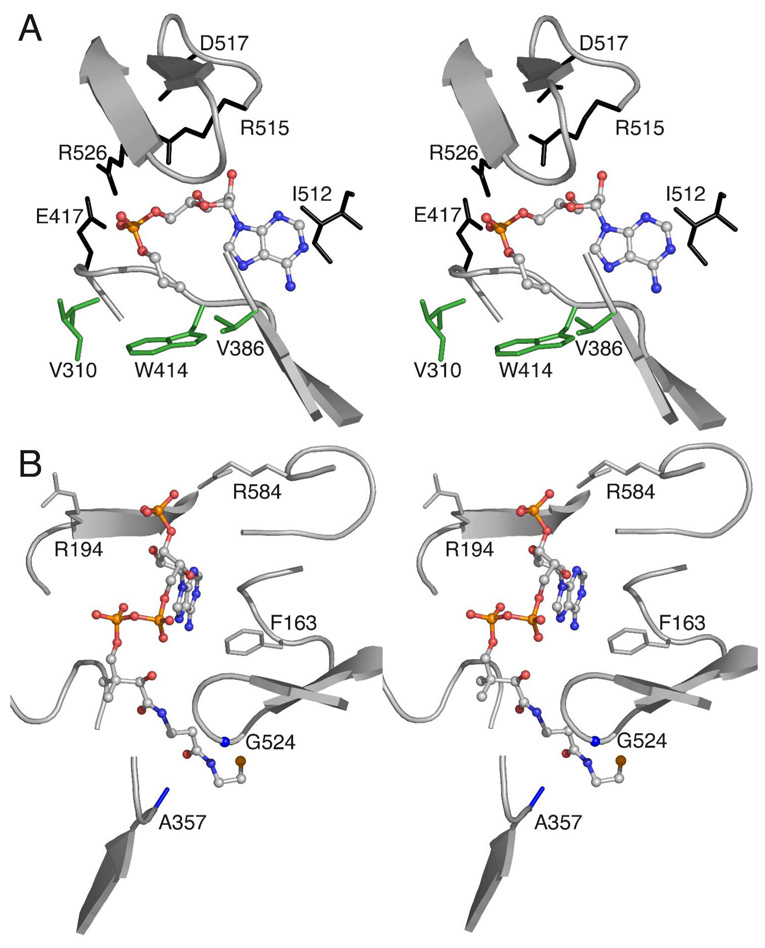
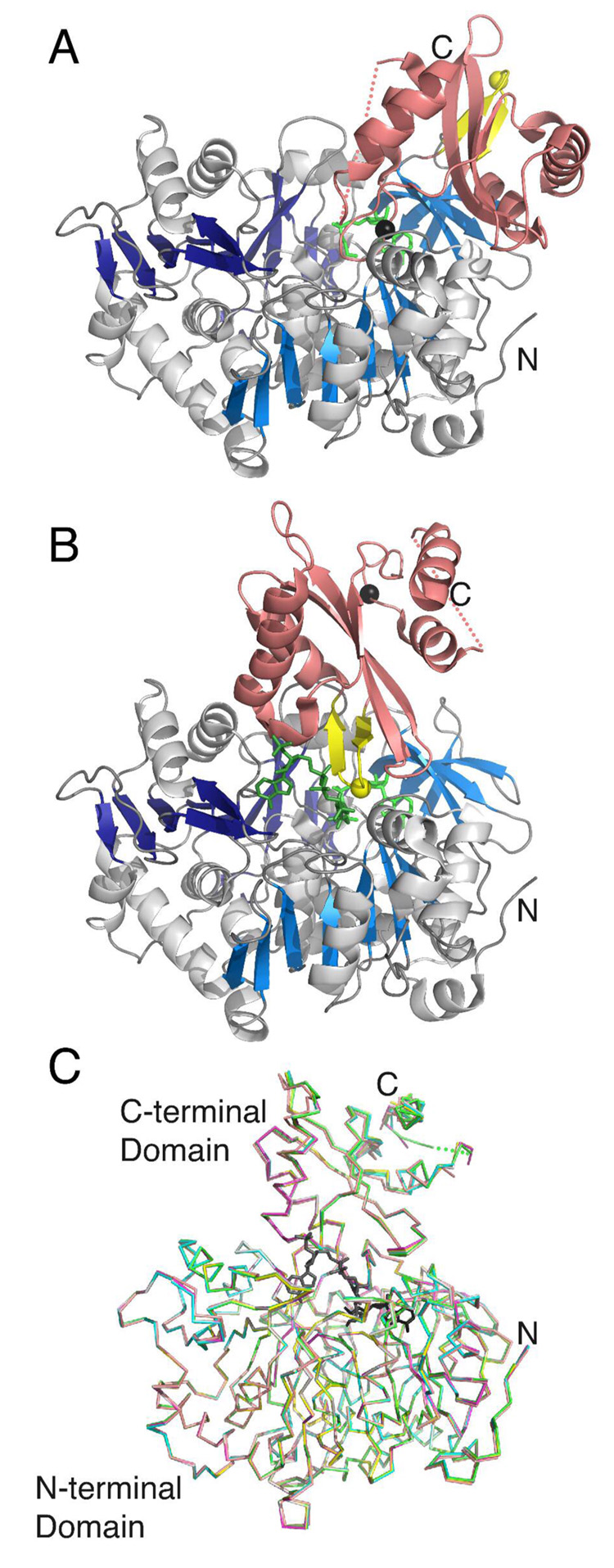
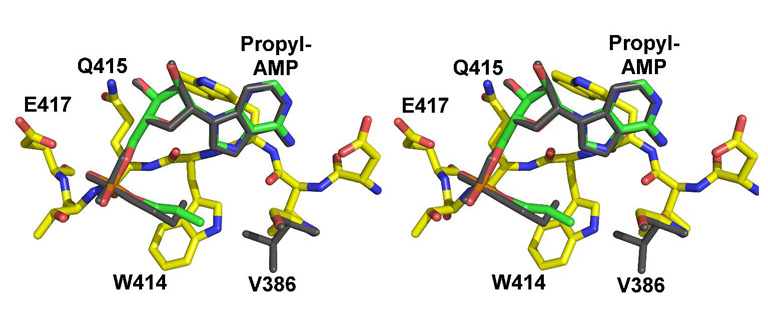
The adenylate-forming enzymes, including acyl-CoA synthetases, the adenylation domains of non-ribosomal peptide synthetases (NRPS), and firefly luciferase, perform two half-reactions in a ping-pong mechanism. We have proposed a domain alternation mechanism for these enzymes whereby, upon completion of the initial adenylation reaction, the C-terminal domain of these enzymes undergoes a 140 degrees rotation to perform the second thioester-forming half-reaction. Structural and kinetic data of mutant enzymes support this hypothesis. We present here mutations to Salmonella enterica acetyl-CoA synthetase (Acs) and test the ability of the enzymes to catalyze the complete reaction and the adenylation half-reaction. Substitution of Lys609 with alanine results in an enzyme that is unable to catalyze the adenylate reaction, while the Gly524 to leucine substitution is unable to catalyze the complete reaction yet catalyzes the adenylation half-reaction with activity comparable to the wild-type enzyme. The positions of these two residues, which are located on the mobile C-terminal domain, strongly support the domain alternation hypothesis. We also present steady-state kinetic data of putative substrate-binding residues and demonstrate that no single residue plays a dominant role in dictating CoA binding. We have also created two mutations in the active site to alter the acyl substrate specificity. Finally, the crystallographic structures of wild-type Acs and mutants R194A, R584A, R584E, K609A, and V386A are presented to support the biochemical analysis.
Figures


References
-
- Sieber SA, Marahiel MA. Molecular mechanisms underlying nonribosomal peptide synthesis: approaches to new antibiotics. Chem Rev. 2005;105:715–738. - PubMed
-
- Marahiel MA, Stachelhaus T, Mootz HD. Modular Peptide Synthetases Involved in Nonribosomal Peptide Synthesis. Chem Rev. 1997;97:2651–2674. - PubMed
-
- Babbitt PC, Kenyon GL, Martin BM, Charest H, Slyvestre M, Scholten JD, Chang KH, Liang PH, Dunaway-Mariano D. Ancestry of the 4-chlorobenzoate dehalogenase: analysis of amino acid sequence identities among families of acyl:adenyl ligases, enoyl-CoA hydratases/isomerases, and acyl-CoA thioesterases. Biochemistry. 1992;31:5594–5604. - PubMed
-
- Dunaway-Mariano D, Babbitt PC. On the origins and functions of the enzymes of the 4-chlorobenzoate to 4-hydroxybenzoate converting pathway. Biodegradation. 1994;5:259–276. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
