

Adaptive decision making and value in the anterior cingulate cortex
- PMID: 17499161
- PMCID: PMC2954047
- DOI: 10.1016/j.neuroimage.2007.03.029
Adaptive decision making and value in the anterior cingulate cortex
Abstract
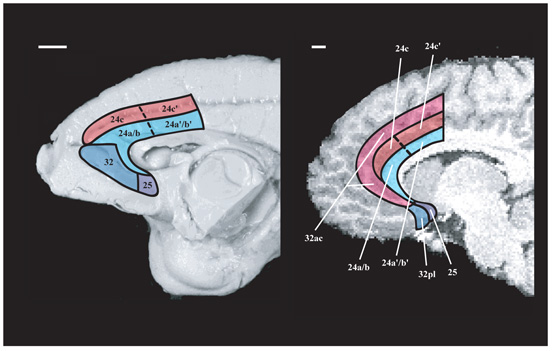
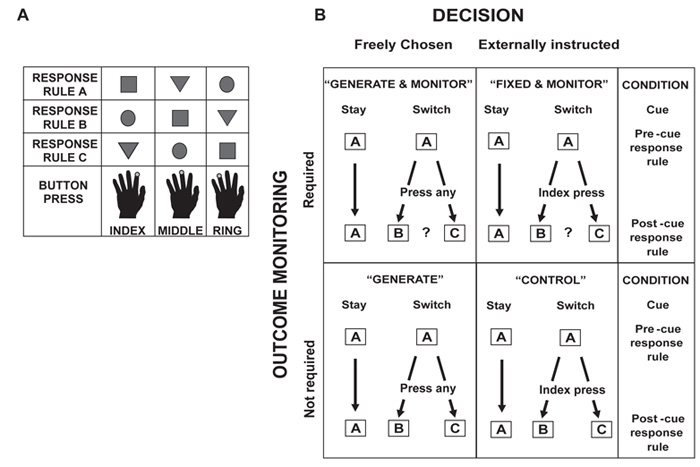
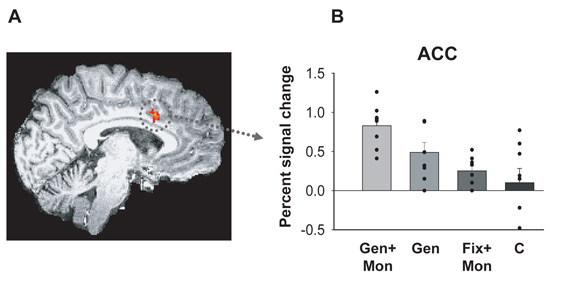
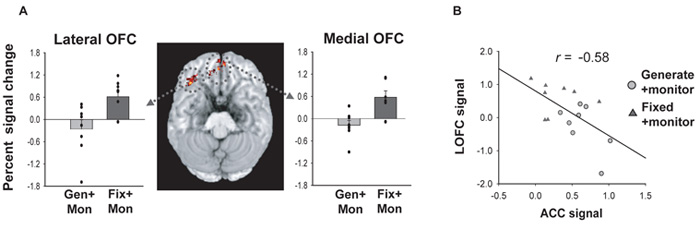
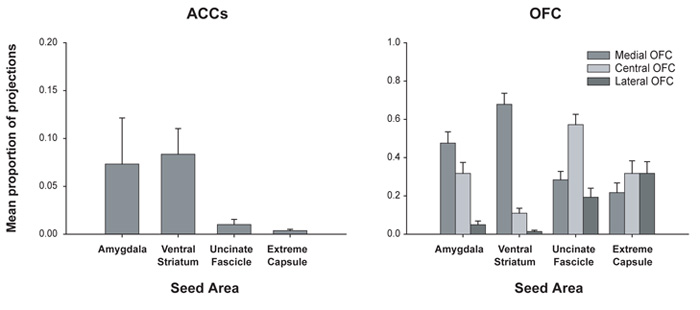
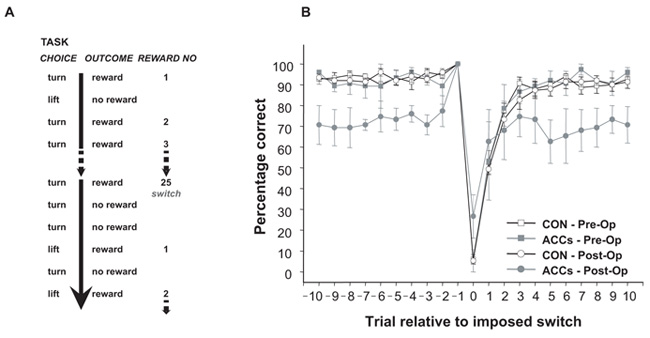
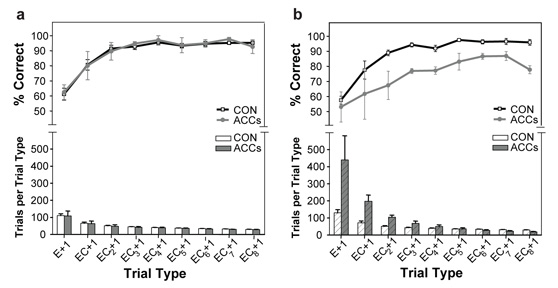
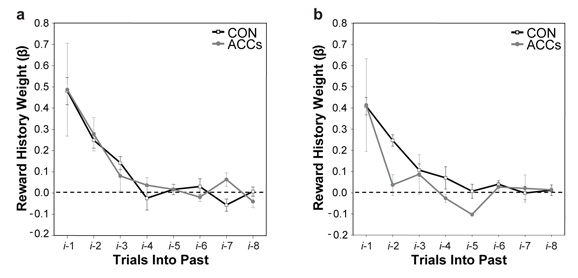
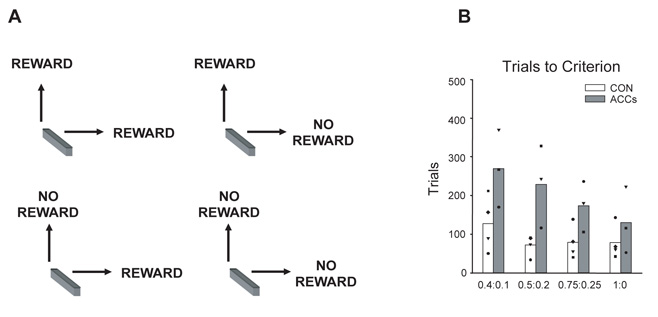
Choosing an appropriate response in an uncertain and varying world is central to adaptive behaviour. The frequent activation of the anterior cingulate cortex (ACC) in a diverse range of tasks has lead to intense interest in and debate over its role in the guidance and control of performance. Here, we consider how this issue can be informed by a series of studies considering the ACC's role in more naturalistic situations where there is no single certain correct response and the relationships between choices and their consequences vary. A neuroimaging study of response switching demonstrates that dorsal ACC is not simply concerned with self-generated responses or error monitoring in isolation, but is instead involved in evaluating the outcome of choices, positive or negative, that have been voluntarily chosen. By contrast, an interconnected part of the orbitofrontal cortex is shown to be more active when attending to consequences of actions instructed by the experimenter. This dissociation is explained with reference to the anatomy of these regions in humans as demonstrated by diffusion weighted imaging. Lesions to a corresponding ACC region in monkeys has no effect on animals' ability to detect or immediately correct errors when response contingencies reverse, but renders them unable to sustain appropriate behaviour due to an impairment in the ability to integrate over time their recent history of choices and outcomes. Taken together, this implies a prominent role for the ACC within a distributed network of regions that determine the dynamic value of actions and guide decision making appropriately.
Figures
References
-
- Allport DA, Styles E-A, Hsieh S. Shifting intentional set: Exploring the dynamic control of tasks. In: Umilta C, editor. Attention and performance series. The MIT Press; Cambridge, MA, US: 1994.
-
- Aston-Jones G, Cohen JD. An integrative theory of locus coeruleus-norepinephrine function: Adaptive gain and optimal performance. Annu Rev Neurosci. 2005;28:403–450. - PubMed
-
- Balleine BW, Dickinson A. Goal-directed instrumental action: Contingency and incentive learning and their cortical substrates. Neuropharmacology. 1998;37:407–419. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
