

Vaccinia virus temperature-sensitive mutants in the A28 gene produce non-infectious virions that bind to cells but are defective in entry
- PMID: 17499330
- PMCID: PMC2062567
- DOI: 10.1016/j.virol.2007.03.060
Vaccinia virus temperature-sensitive mutants in the A28 gene produce non-infectious virions that bind to cells but are defective in entry
Abstract
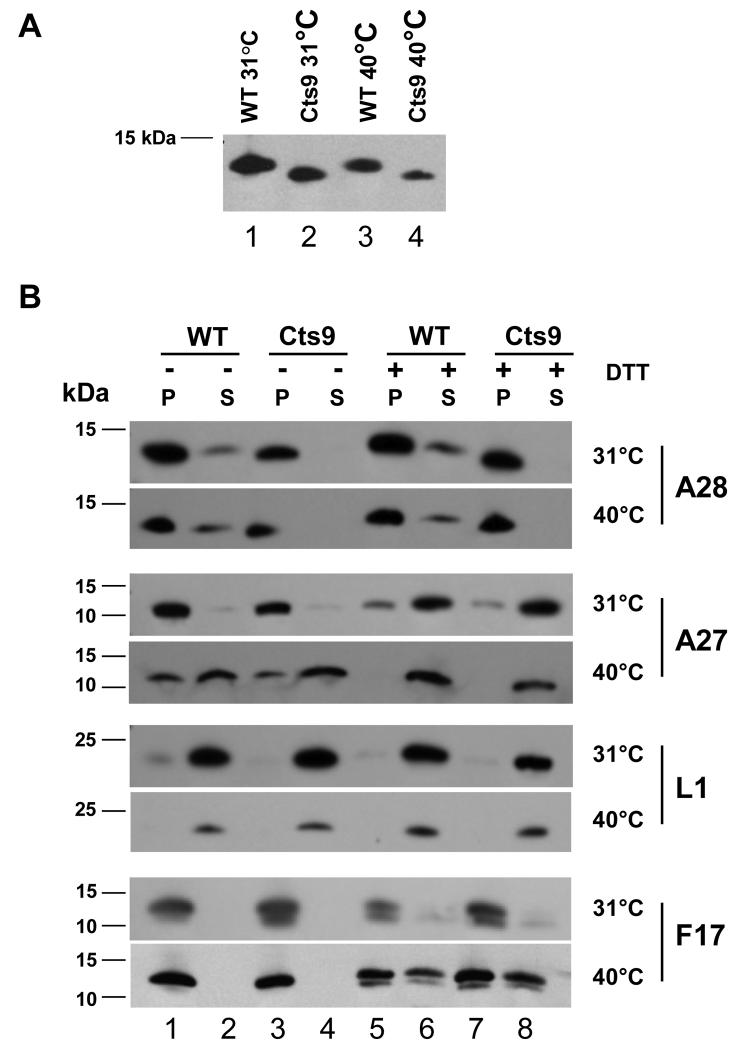
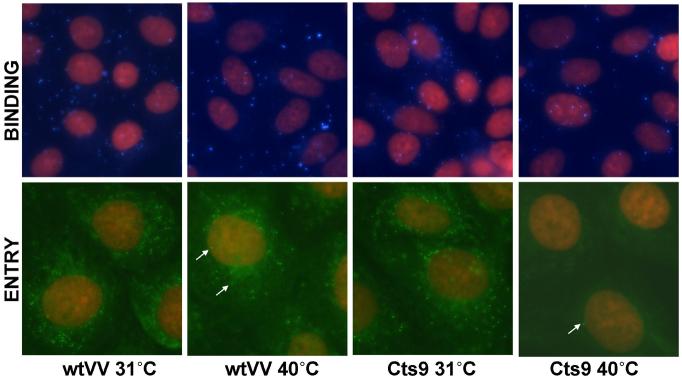
The vaccinia virus temperature-sensitive mutations Cts6 and Cts9 were mapped by marker rescue and DNA sequencing to the A28 gene. Cts6 and Cts9 contain an identical 2-bp deletion truncating the A28 protein and removing the fourth conserved cysteine near the C-terminus. Cts9 mutant virions produced at 40 degrees C were non-infectious and unable to cause cytopathic effect. However, the mutant A28 protein localized to purified mature virions (MV) at 31 degrees C and 40 degrees C. MV of Cts9 produced at 40 degrees C bound to cells but did not enter cells. Low pH treatment of Cts9-infected cells at 18 h p.i. failed to produce fusion from within at 40 degrees C, but gave fusion at 31 degrees C. Adsorption of Cts9 mutant virions to cells followed by low pH treatment showed a defect in fusion from without. The Cts9 phenotype suggests that the A28 protein is involved in both virus entry and cell-cell fusion, and supports the linkage between the two processes.
Figures





References
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current Protocols in Molecular Biology. John Wiley and Sons, Inc.; New York: 1994.
-
- Carter GC, Law M, Hollinshead M, Smith GL. Entry of the vaccinia virus intracellular mature virion and its interactions with glycosaminoglycans. J Gen.Virol. 2005;86:1279–1290. - PubMed
-
- Condit RC, Motyczka A. Isolation and preliminary characterization of temperature-sensitive mutants of vaccinia virus. Virology. 1981;113:224–241. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
