

Integration of estrogen and Wnt signaling circuits by the polycomb group protein EZH2 in breast cancer cells
- PMID: 17502350
- PMCID: PMC1951944
- DOI: 10.1128/MCB.00162-07
Integration of estrogen and Wnt signaling circuits by the polycomb group protein EZH2 in breast cancer cells
Abstract
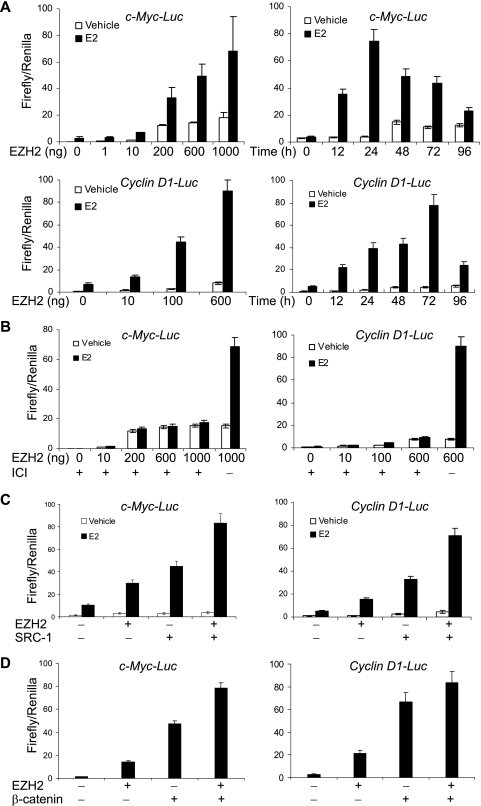
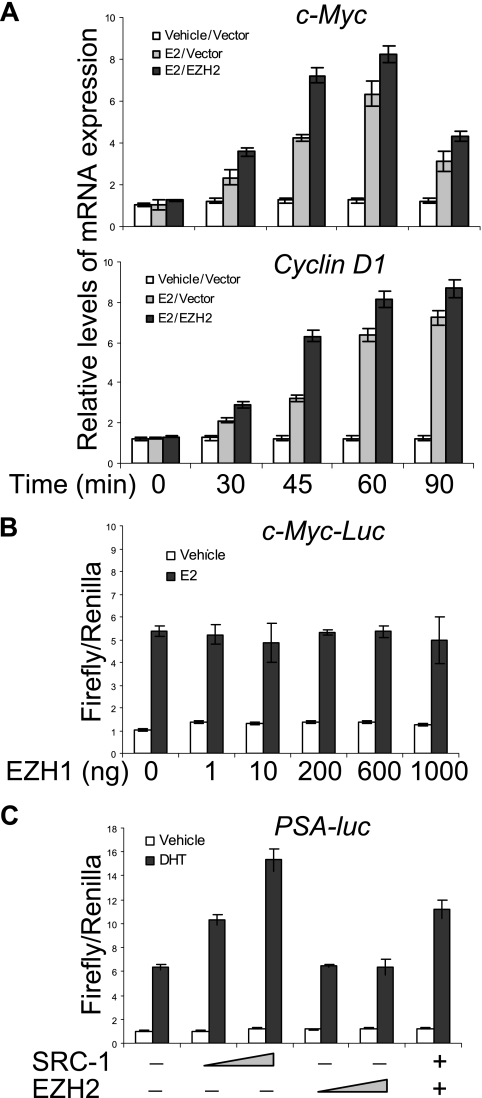
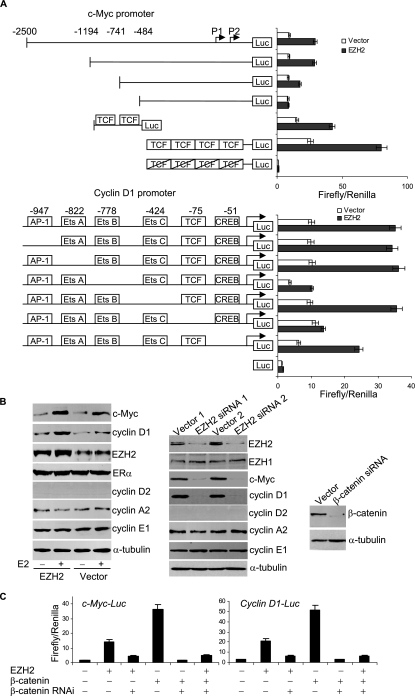
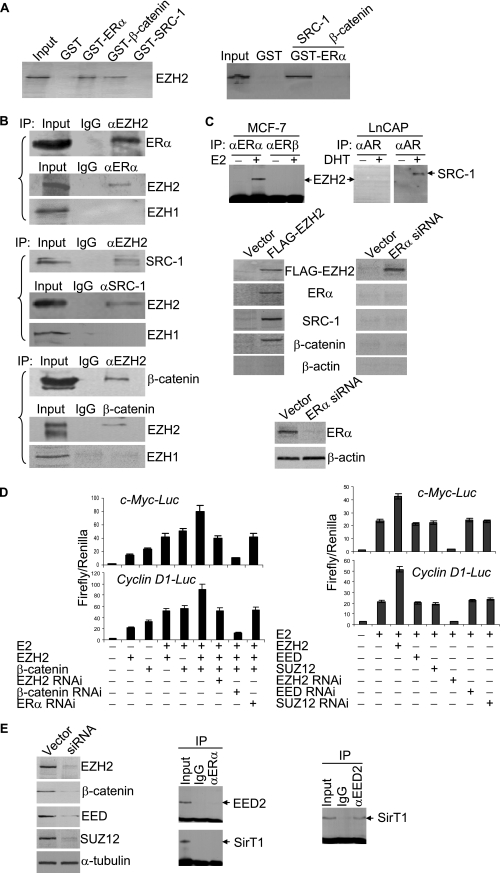
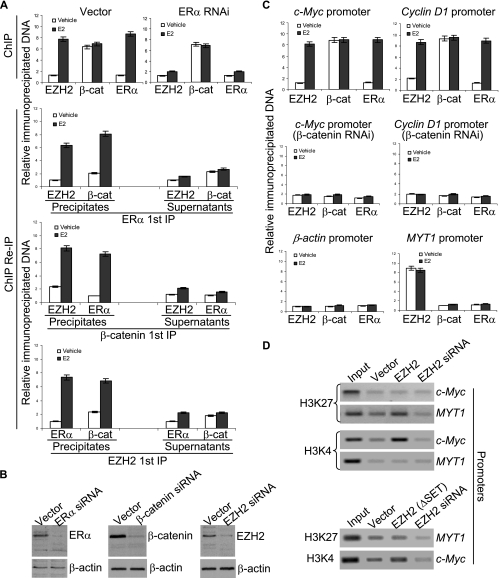
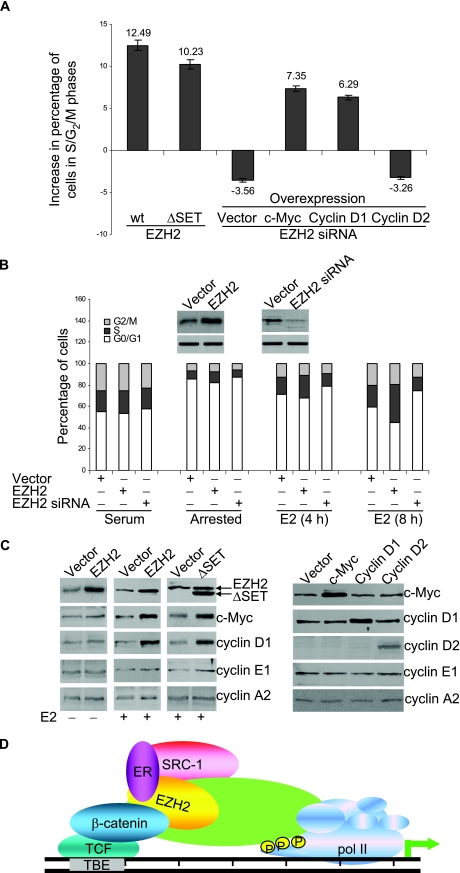
Essential for embryonic development, the polycomb group protein enhancer of zeste homolog 2 (EZH2) is overexpressed in breast and prostate cancers and is implicated in the growth and aggression of the tumors. The tumorigenic mechanism underlying EZH2 overexpression is largely unknown. It is believed that EZH2 exerts its biological activity as a transcription repressor. However, we report here that EZH2 functions in gene transcriptional activation in breast cancer cells. We show that EZH2 transactivates genes that are commonly targeted by estrogen and Wnt signaling pathways. We demonstrated that EZH2 physically interacts directly with estrogen receptor alpha and beta-catenin, thus connecting the estrogen and Wnt signaling circuitries, functionally enhances gene transactivation by estrogen and Wnt pathways, and phenotypically promotes cell cycle progression. In addition, we identified the transactivation activity of EZH2 in its two N-terminal domains and demonstrated that these structures serve as platforms to connect transcription factors and the Mediator complex. Our experiments indicated that EZH2 is a dual function transcription regulator with a dynamic activity, and we provide a mechanism for EZH2 in tumorigenesis.
Figures

Similar articles
-
EZH2-mediated concordant repression of Wnt antagonists promotes β-catenin-dependent hepatocarcinogenesis.Cancer Res. 2011 Jun 1;71(11):4028-39. doi: 10.1158/0008-5472.CAN-10-3342. Epub 2011 Apr 21. Cancer Res. 2011. PMID: 21512140
-
EZH2 regulates the transcription of estrogen-responsive genes through association with REA, an estrogen receptor corepressor.Breast Cancer Res Treat. 2008 Jan;107(2):235-42. doi: 10.1007/s10549-007-9542-7. Epub 2007 Apr 24. Breast Cancer Res Treat. 2008. PMID: 17453341
-
MEK-ERK pathway regulates EZH2 overexpression in association with aggressive breast cancer subtypes.Oncogene. 2011 Sep 29;30(39):4118-28. doi: 10.1038/onc.2011.118. Epub 2011 Apr 18. Oncogene. 2011. PMID: 21499305
-
The Polycomb group protein Enhancer of Zeste 2: its links to DNA repair and breast cancer.J Mol Histol. 2006 Sep;37(5-7):219-23. doi: 10.1007/s10735-006-9042-9. Epub 2006 Jul 20. J Mol Histol. 2006. PMID: 16855786 Review.
-
Importance of Ezh2 polycomb protein in tumorigenesis process interfering with the pathway of growth suppressive key elements.J Cell Physiol. 2008 Feb;214(2):295-300. doi: 10.1002/jcp.21241. J Cell Physiol. 2008. PMID: 17786943 Review.
Cited by
-
Polycomb Repressive Complex 2: a Dimmer Switch of Gene Regulation in Calvarial Bone Development.Curr Osteoporos Rep. 2020 Aug;18(4):378-387. doi: 10.1007/s11914-020-00603-5. Curr Osteoporos Rep. 2020. PMID: 32748325 Free PMC article. Review.
-
PRC2 during vertebrate organogenesis: a complex in transition.Dev Biol. 2012 Jul 15;367(2):91-9. doi: 10.1016/j.ydbio.2012.04.030. Epub 2012 May 4. Dev Biol. 2012. PMID: 22565092 Free PMC article. Review.
-
LncRNA MALAT1 facilitates inflammasome activation via epigenetic suppression of Nrf2 in Parkinson's disease.Mol Brain. 2020 Sep 24;13(1):130. doi: 10.1186/s13041-020-00656-8. Mol Brain. 2020. PMID: 32972446 Free PMC article.
-
Single-Cell Sequencing Reveals the Novel Role of Ezh2 in NK Cell Maturation and Function.Front Immunol. 2021 Oct 26;12:724276. doi: 10.3389/fimmu.2021.724276. eCollection 2021. Front Immunol. 2021. PMID: 34764950 Free PMC article.
-
Epigenetic reprogramming of HOXC10 in endocrine-resistant breast cancer.Sci Transl Med. 2014 Mar 26;6(229):229ra41. doi: 10.1126/scitranslmed.3008326. Sci Transl Med. 2014. PMID: 24670685 Free PMC article.
References
-
- Acconcia, F., B. Manavathi, J. Mascarenhas, A. H. Talukder, G. Mills, and R. Kumar. 2006. An inherent role of integrin-linked kinase-estrogen receptor alpha interaction in cell migration. Cancer Res. 66:11030-11038. - PubMed
-
- Attwooll, C., S. Oddi, P. Cartwright, E. Prosperini, K. Agger, P. Steensgaard, C. Wagener, C. Sardet, M. C. Moroni, and K. Helin. 2005. A novel repressive E2F6 complex containing the polycomb group protein, EPC1, that interacts with EZH2 in a proliferation-specific manner. J. Biol. Chem. 280:1199-1208. - PubMed
-
- Cao, R., L. Wang, H. Wang, L. Xia, H. Erdjument-Bromage, P. Tempst, R. S. Jones, and Y. Zhang. 2002. Role of histone H3 lysine 27 methylation in Polycomb-group silencing. Science 298:1039-1043. - PubMed
-
- Cao, R., and Y. Zhang. 2004. The functions of E(Z)/EZH2-mediated methylation of lysine 27 in histone H3. Curr. Opin. Genet. Dev. 14:155-164. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical