

Eca1, a sarcoplasmic/endoplasmic reticulum Ca2+-ATPase, is involved in stress tolerance and virulence in Cryptococcus neoformans
- PMID: 17502401
- PMCID: PMC1932933
- DOI: 10.1128/IAI.01977-06
Eca1, a sarcoplasmic/endoplasmic reticulum Ca2+-ATPase, is involved in stress tolerance and virulence in Cryptococcus neoformans
Abstract
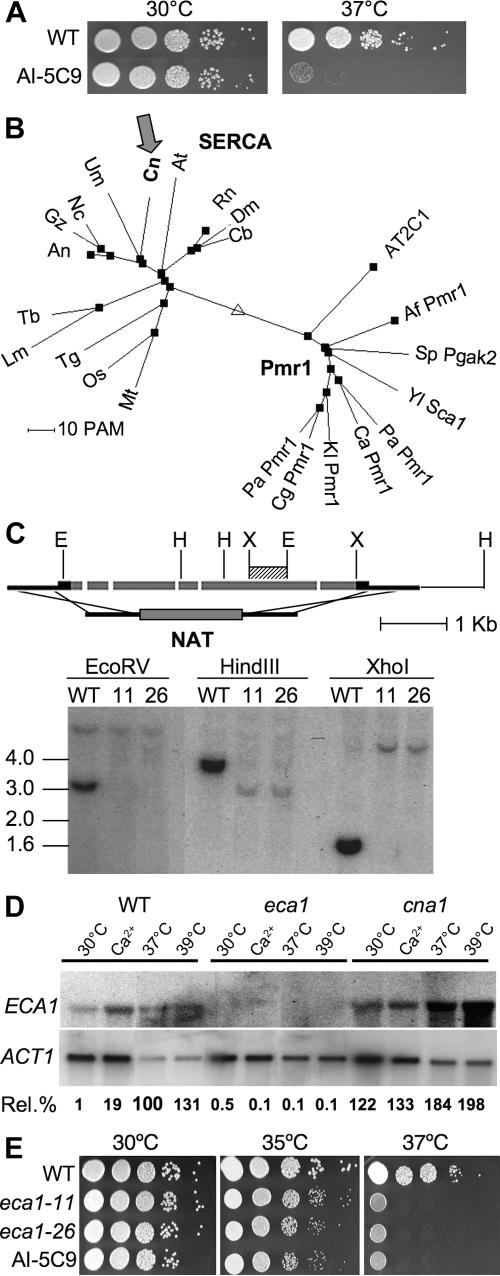
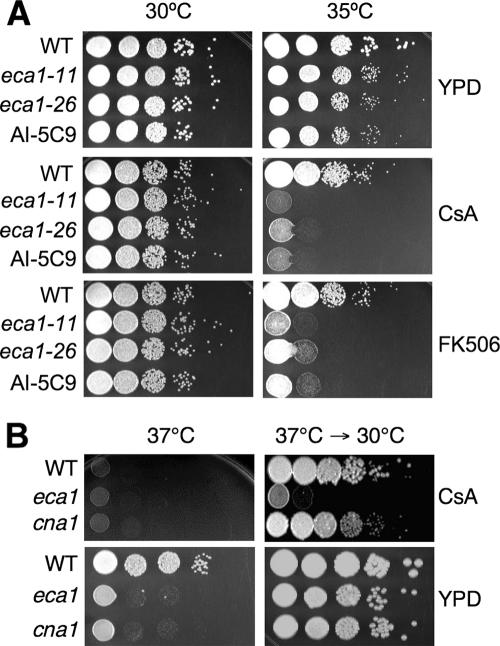
The basidiomycetous fungal pathogen Cryptococcus neoformans is adapted to survive challenges in the soil and environment and within the unique setting of the mammalian host. A C. neoformans mutant was isolated with enhanced virulence in a soil amoeba model that nevertheless exhibits dramatically reduced growth at mammalian body temperature (37 degrees C). This mutant phenotype results from an insertion in the ECA1 gene, which encodes a sarcoplasmic/endoplasmic reticulum (ER) Ca2+-ATPase (SERCA)-type calcium pump. Infection in murine macrophages, amoebae (Acanthamoeba castellanii), nematodes (Caenorhabditis elegans), and wax moth (Galleria mellonella) larvae revealed that the eca1 mutants are virulent or hypervirulent at permissive growth temperatures but attenuated at 37 degrees C. Deletion mutants lacking the entire ECA1 gene were also hypersensitive to the calcineurin inhibitors cyclosporin and FK506 and to ER and osmotic stresses. An eca1Delta cna1Delta mutant lacking both Eca1 and the calcineurin catalytic subunit was more sensitive to high temperature and ER stresses than the single mutants and exhibited reduced survival in C. elegans and attenuated virulence towards wax moth larvae at temperatures that permit normal growth in vitro. Eca1 is likely involved in maintaining ER function, thus contributing to stress tolerance and virulence acting in parallel with Ca2+-calcineurin signaling.
Figures






References
-
- Alspaugh, J. A., L. M. Cavallo, J. R. Perfect, and J. Heitman. 2000. RAS1 regulates filamentation, mating and growth at high temperature of Cryptococcus neoformans. Mol. Microbiol. 36:352-365. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
