

Evolution of robustness to noise and mutation in gene expression dynamics
- PMID: 17502916
- PMCID: PMC1855988
- DOI: 10.1371/journal.pone.0000434
Evolution of robustness to noise and mutation in gene expression dynamics
Abstract
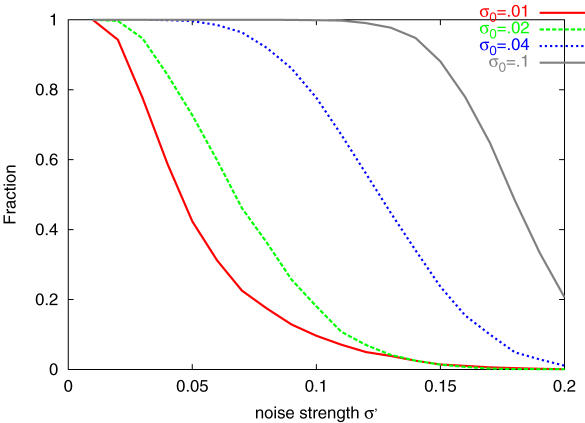
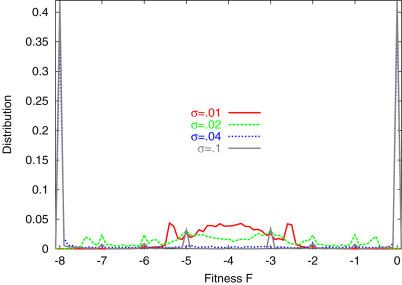
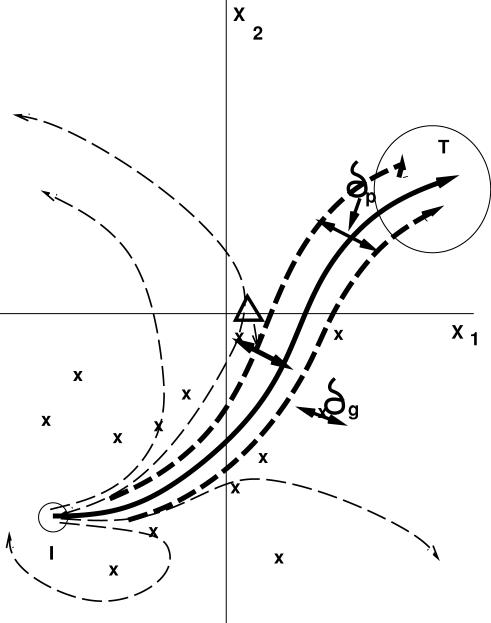
Phenotype of biological systems needs to be robust against mutation in order to sustain themselves between generations. On the other hand, phenotype of an individual also needs to be robust against fluctuations of both internal and external origins that are encountered during growth and development. Is there a relationship between these two types of robustness, one during a single generation and the other during evolution? Could stochasticity in gene expression have any relevance to the evolution of these types of robustness? Robustness can be defined by the sharpness of the distribution of phenotype; the variance of phenotype distribution due to genetic variation gives a measure of 'genetic robustness', while that of isogenic individuals gives a measure of 'developmental robustness'. Through simulations of a simple stochastic gene expression network that undergoes mutation and selection, we show that in order for the network to acquire both types of robustness, the phenotypic variance induced by mutations must be smaller than that observed in an isogenic population. As the latter originates from noise in gene expression, this signifies that the genetic robustness evolves only when the noise strength in gene expression is larger than some threshold. In such a case, the two variances decrease throughout the evolutionary time course, indicating increase in robustness. The results reveal how noise that cells encounter during growth and development shapes networks' robustness to stochasticity in gene expression, which in turn shapes networks' robustness to mutation. The necessary condition for evolution of robustness, as well as the relationship between genetic and developmental robustness, is derived quantitatively through the variance of phenotypic fluctuations, which are directly measurable experimentally.
Conflict of interest statement
Figures

Similar articles
-
Phenotypic plasticity and robustness: evolutionary stability theory, gene expression dynamics model, and laboratory experiments.Adv Exp Med Biol. 2012;751:249-78. doi: 10.1007/978-1-4614-3567-9_12. Adv Exp Med Biol. 2012. PMID: 22821462
-
Relevance of phenotypic noise to adaptation and evolution.IET Syst Biol. 2008 Sep;2(5):234-46. doi: 10.1049/iet-syb:20070078. IET Syst Biol. 2008. PMID: 19045819
-
Evolution of phenotypic fluctuation under host-parasite interactions.PLoS Comput Biol. 2021 Nov 9;17(11):e1008694. doi: 10.1371/journal.pcbi.1008694. eCollection 2021 Nov. PLoS Comput Biol. 2021. PMID: 34752445 Free PMC article.
-
The Evolution of Variability and Robustness in Neural Development.Trends Neurosci. 2018 Sep;41(9):577-586. doi: 10.1016/j.tins.2018.05.007. Epub 2018 Jun 4. Trends Neurosci. 2018. PMID: 29880259 Review.
-
Links between evolutionary processes and phenotypic robustness in microbes.Semin Cell Dev Biol. 2019 Apr;88:46-53. doi: 10.1016/j.semcdb.2018.05.017. Epub 2018 Jun 19. Semin Cell Dev Biol. 2019. PMID: 29803630 Review.
Cited by
-
Pervasive robustness in biological systems.Nat Rev Genet. 2015 Aug;16(8):483-96. doi: 10.1038/nrg3949. Nat Rev Genet. 2015. PMID: 26184598 Review.
-
Evolutionary dynamics, evolutionary forces, and robustness: A nonequilibrium statistical mechanics perspective.Proc Natl Acad Sci U S A. 2022 Mar 29;119(13):e2112083119. doi: 10.1073/pnas.2112083119. Epub 2022 Mar 21. Proc Natl Acad Sci U S A. 2022. PMID: 35312370 Free PMC article.
-
An Approach for Systems-Level Understanding of Prostate Cancer from High-Throughput Data Integration to Pathway Modeling and Simulation.Cells. 2022 Dec 19;11(24):4121. doi: 10.3390/cells11244121. Cells. 2022. PMID: 36552885 Free PMC article.
-
Developmental robustness by obligate interaction of class B floral homeotic genes and proteins.PLoS Comput Biol. 2009 Jan;5(1):e1000264. doi: 10.1371/journal.pcbi.1000264. Epub 2009 Jan 16. PLoS Comput Biol. 2009. PMID: 19148269 Free PMC article.
-
Evolution-development congruence in pattern formation dynamics: Bifurcations in gene expression and regulation of networks structures.J Exp Zool B Mol Dev Evol. 2016 Jan;326(1):61-84. doi: 10.1002/jez.b.22666. J Exp Zool B Mol Dev Evol. 2016. PMID: 26678220 Free PMC article.
References
-
- Barkai N, Leibler S. Robustness in simple biochemical networks. Nature. 1997:913–917. - PubMed
-
- Alon U, Surette MG, Barkai N, Leibler S. Robustness in bacterial chemotaxis, Nature. 1999:168–171. - PubMed
-
- Wagner A. Robustness against mutations in genetic networks of yeast. Nature Genetics, 2000:355–361. - PubMed
-
- de Visser JAGM, et al. Evolution and detection of genetic robustness. Evolution. 2003:1959–1972. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
