

A mutation in CCDC50, a gene encoding an effector of epidermal growth factor-mediated cell signaling, causes progressive hearing loss
- PMID: 17503326
- PMCID: PMC1867096
- DOI: 10.1086/518311
A mutation in CCDC50, a gene encoding an effector of epidermal growth factor-mediated cell signaling, causes progressive hearing loss
Abstract
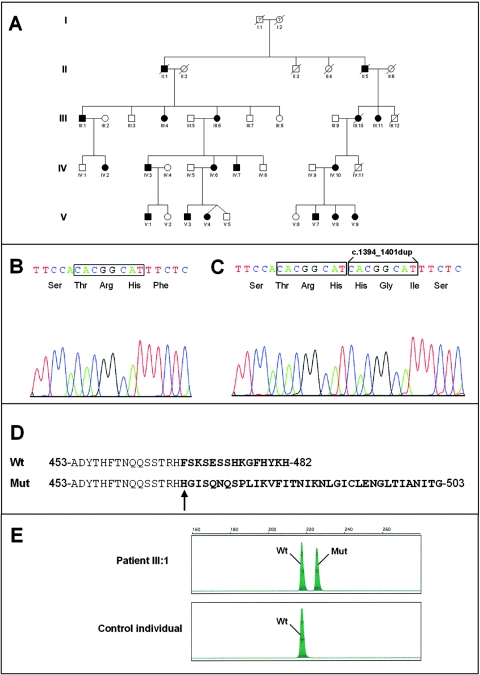
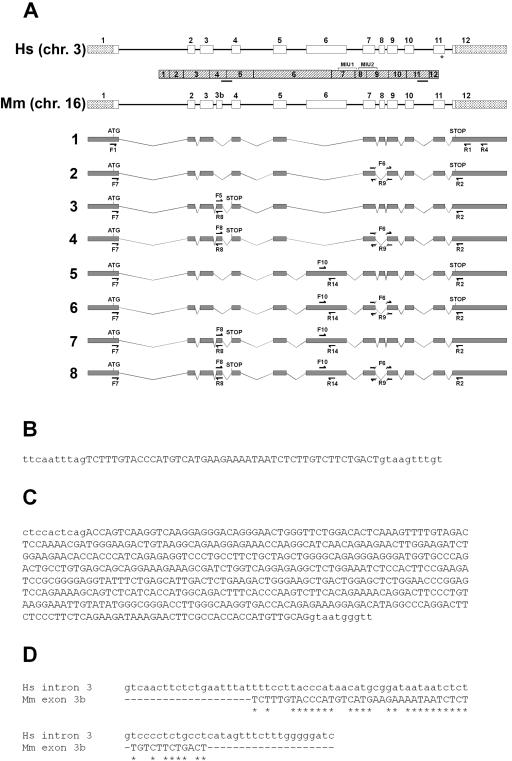
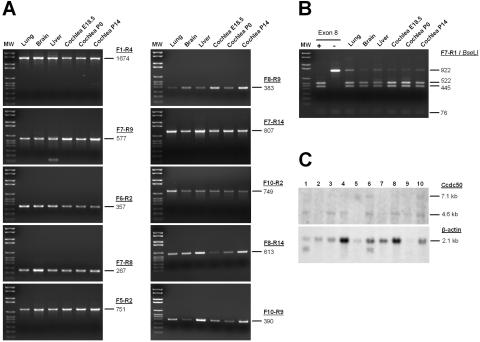
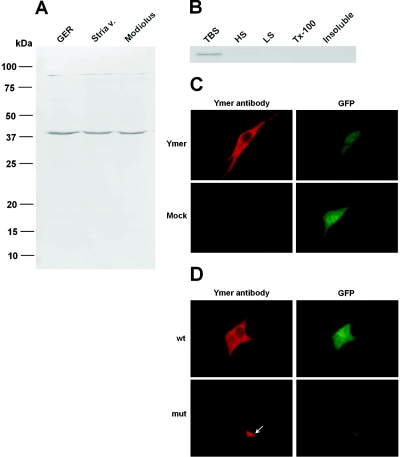
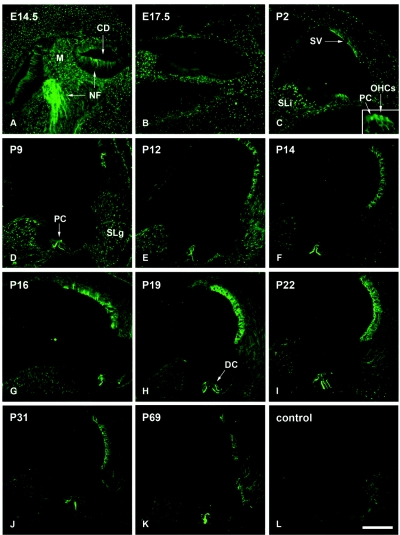
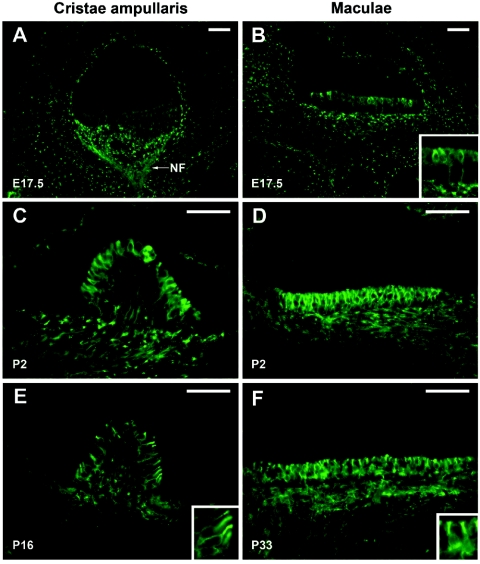
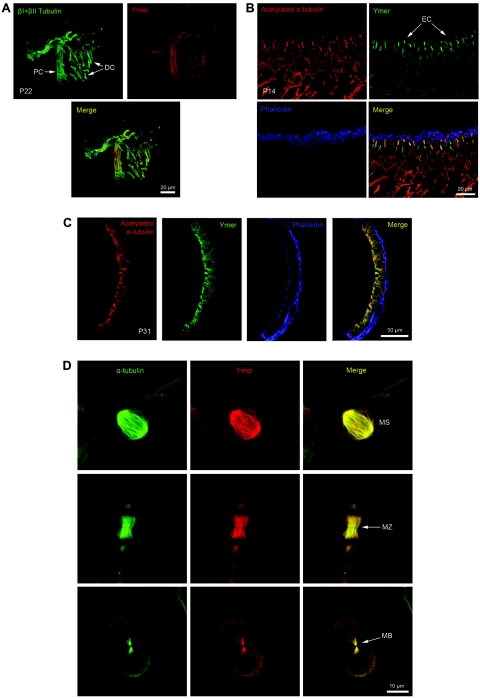
We previously mapped a novel autosomal dominant deafness locus, DFNA44, by studying a family with postlingual, progressive, nonsyndromic hearing loss. We report here on the identification of a mutation in CCDC50 as the cause of hearing loss in the family. CCDC50 encodes Ymer, an effector of epidermal growth factor (EGF)-mediated cell signaling that is ubiquitously expressed in different organs and has been suggested to inhibit down-regulation of the EGF receptor. We have examined its expression pattern in mouse inner ear. Western blotting and cell transfection results indicate that Ymer is a soluble, cytoplasmic protein, and immunostaining shows that Ymer is expressed in a complex spatiotemporal pattern during inner ear development. In adult inner ear, the expression of Ymer is restricted to the pillar cells of the cochlea, the stria vascularis, and the vestibular sensory epithelia, where it shows spatial overlap with the microtubule-based cytoskeleton. In dividing cells, Ymer colocalizes with microtubules of the mitotic apparatus. We suggest that DFNA44 hearing loss may result from a time-dependent disorganization of the microtubule-based cytoskeleton in the pillar cells and stria vascularis of the adult auditory system.
Figures
References
Web Resources
-
- ClustalW, http://www.ebi.ac.uk/clustalw
-
- GenBank, http://www.ncbi.nlm.nih.gov/GenBank/ (for CCDC50 transcripts 1 [accession number NM_174908] and 2 [accession number NM_178335], Ccdc50 transcripts 1 [accession number NM_026202.2] and 2 [accession number NM_001025615.1]), mouse organ of Corti [accession number BQ567072.1] and adult mouse testis [accession number AK016827.1] Ccdc50 ESTs, and Ccdc50 pseudogene [accession number NG_005190.2])
-
- Hereditary Hearing Loss Homepage, http://webhost.ua.ac.be/hhh/
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
