

Sequential regulation of diacylglycerol acyltransferase 2 expression by CAAT/enhancer-binding protein beta (C/EBPbeta) and C/EBPalpha during adipogenesis
- PMID: 17504763
- PMCID: PMC2254492
- DOI: 10.1074/jbc.M702871200
Sequential regulation of diacylglycerol acyltransferase 2 expression by CAAT/enhancer-binding protein beta (C/EBPbeta) and C/EBPalpha during adipogenesis
Abstract
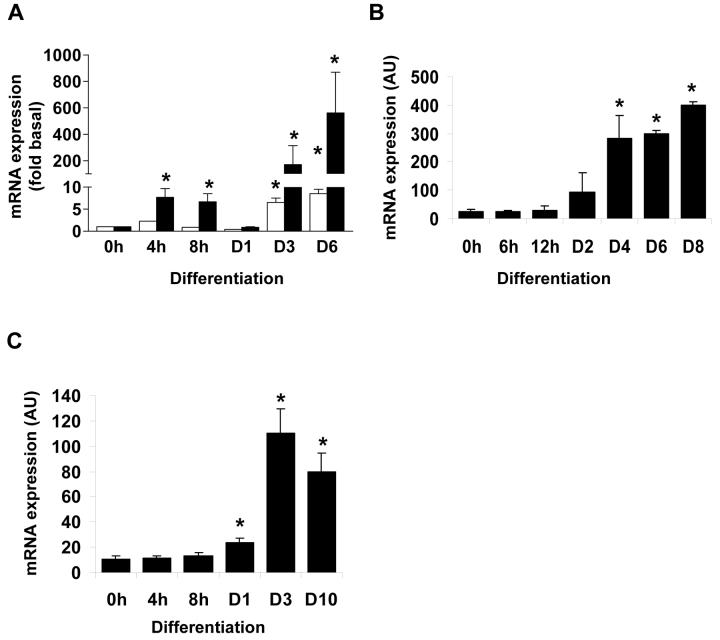
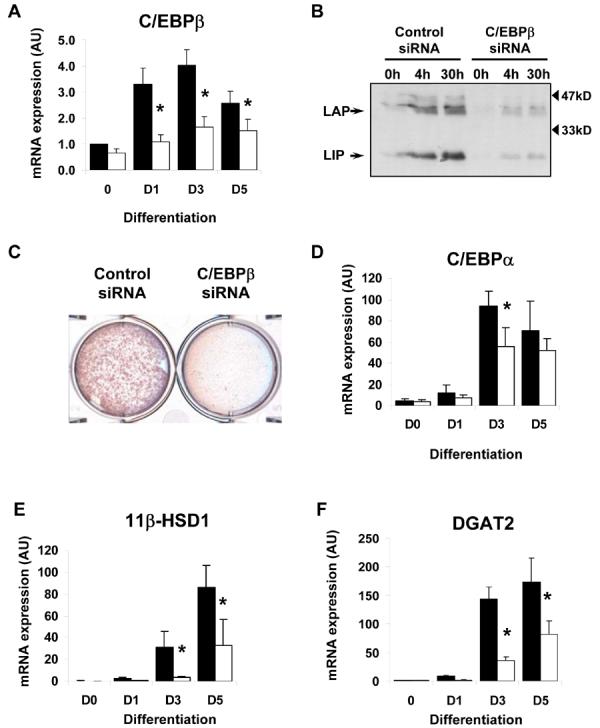
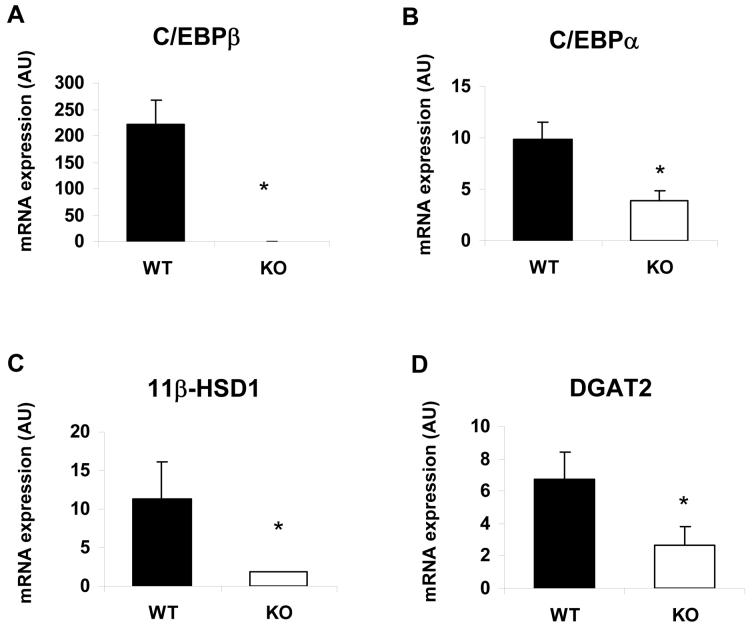
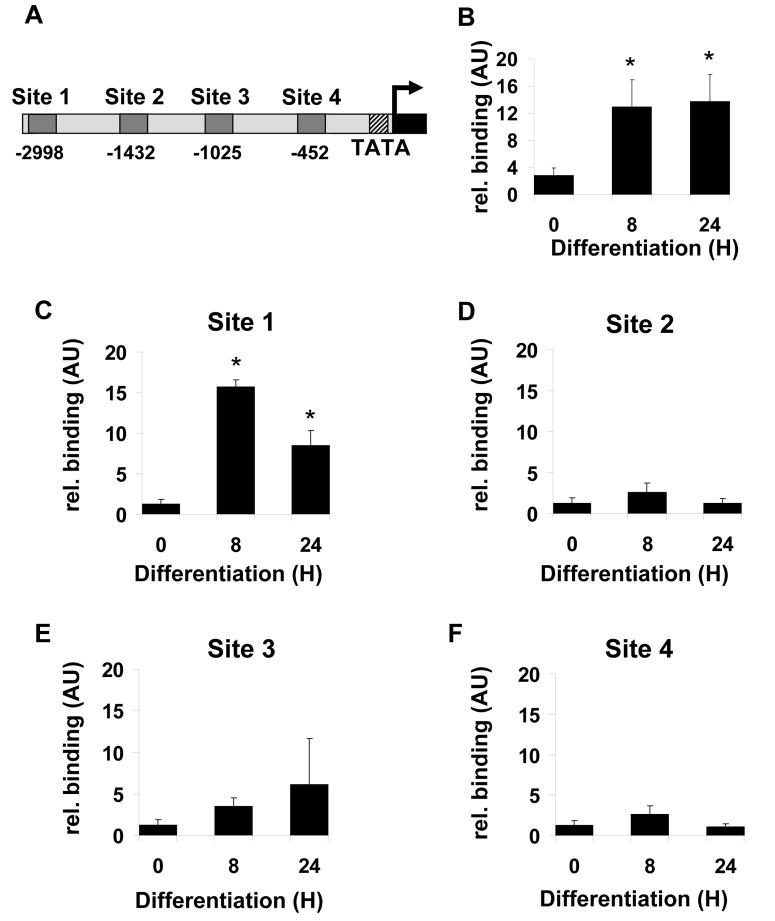
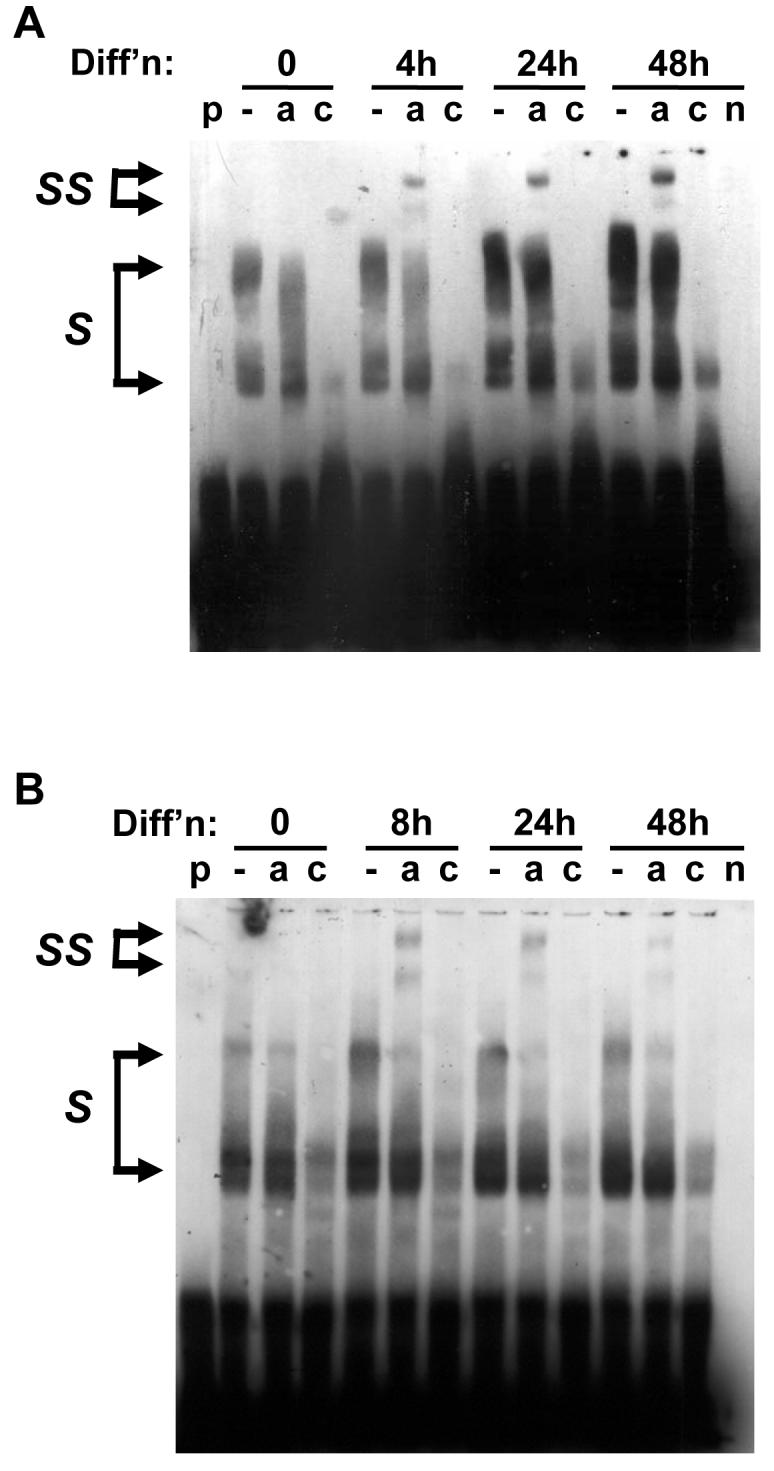
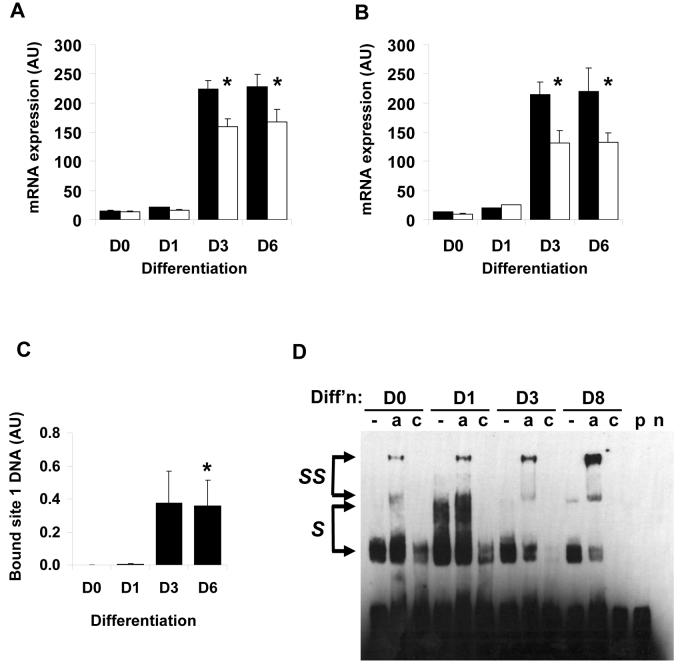
Diacylglycerol acyltransferase 2 (DGAT2) catalyzes the final step of triacylglycerol (TG) synthesis. Despite the existence of an alternative acyltransferase (DGAT1), mice lacking DGAT2 have a severe deficiency of TG in adipose tissue, indicating a nonredundant role for this enzyme in adipocyte TG synthesis. We have studied the regulation of DGAT2 expression during adipogenesis. In both isolated murine preadipocytes and 3T3-L1 cells the temporal pattern of DGAT2 expression closely mimicked that of genes whose expression is regulated by CAAT/enhancer-binding protein beta (C/EBPbeta). Inhibition of C/EBPbeta expression in differentiating preadipocytes reduced DGAT2 expression, and electrophoretic mobility shift assay and chromatin immunoprecipitation experiments identified a promoter element in the DGAT2 gene that is likely to mediate this effect. The importance of C/EBPbeta in adipocyte expression of DGAT2 was confirmed by the finding of reduced DGAT2 expression in the adipose tissue of C/EBPbeta-null animals. However, DGAT2 expression is maintained at high levels during the later stages of adipogenesis, when C/EBPbeta levels decline. We show that, at these later stages of differentiation, C/EBPalpha is capable of substituting for C/EBPbeta at the same promoter element. These observations provide novel insight into the transcriptional regulation of DGAT2 expression. Moreover, they further refine the complex and serial roles of the C/EBP family of transcription factors in inducing and maintaining the metabolic properties of mature adipocytes.
Figures



References
-
- Camp HS, Ren D, Leff T. Trends Mol Med. 2002;8(9):442–447. - PubMed
-
- Agarwal AK, Arioglu E, De Almeida S, Akkoc N, Taylor SI, Bowcock AM, Barnes RI, Garg A. Nat Genet. 2002;31(1):21–23. - PubMed
-
- Magre J, Delepine M, Van Maldergem L, Robert JJ, Maassen JA, Meier M, Panz VR, Kim CA, Tubiana-Rufi N, Czernichow P, Seemanova E, Buchanan CR, Lacombe D, Vigouroux C, Lascols O, Kahn CR, Capeau J, Lathrop M. Diabetes. 2003;52(6):1573–1578. - PubMed
-
- Smith SJ, Cases S, Jensen DR, Chen HC, Sande E, Tow B, Sanan DA, Raber J, Eckel RH, Farese RV., Jr. Nat Genet. 2000;25(1):87–90. - PubMed
-
- Buhman KK, Smith SJ, Stone SJ, Repa JJ, Wong JS, Knapp FF, Jr., Burri BJ, Hamilton RL, Abumrad NA, Farese RV., Jr. J Biol Chem. 2002;277(28):25474–25479. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
