

A global view of gene expression in lithium and zinc treated sea urchin embryos: new components of gene regulatory networks
- PMID: 17506889
- PMCID: PMC1929154
- DOI: 10.1186/gb-2007-8-5-r85
A global view of gene expression in lithium and zinc treated sea urchin embryos: new components of gene regulatory networks
Abstract
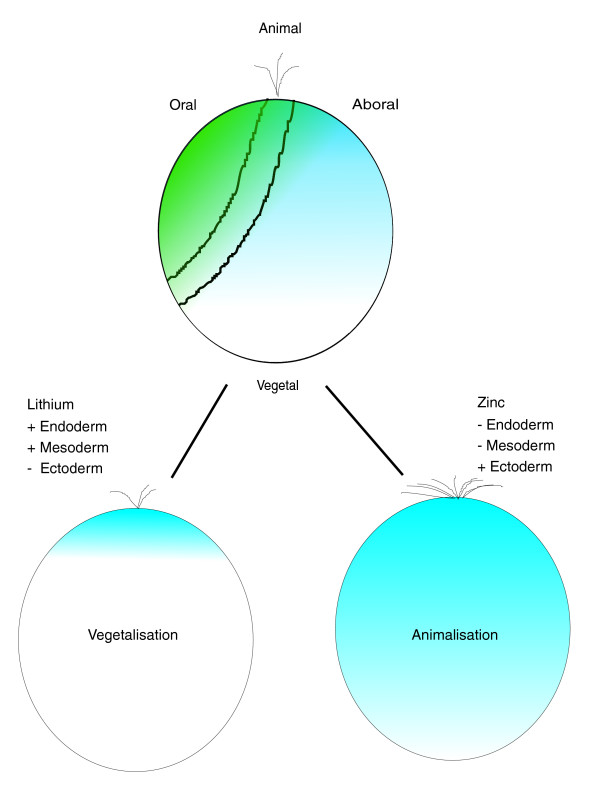
Background: The genome of the sea urchin Strongylocentrotus purpuratus has recently been sequenced because it is a major model system for the study of gene regulatory networks. Embryonic expression patterns for most genes are unknown, however.
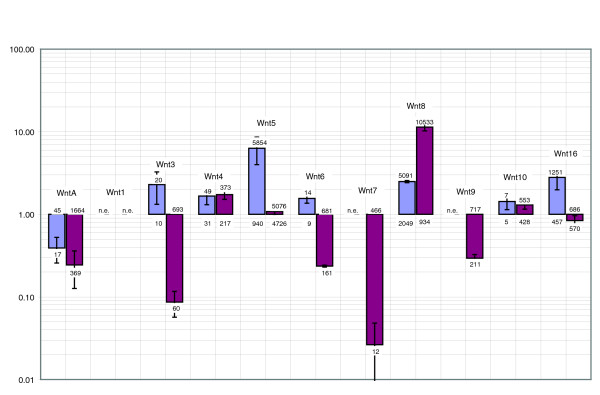
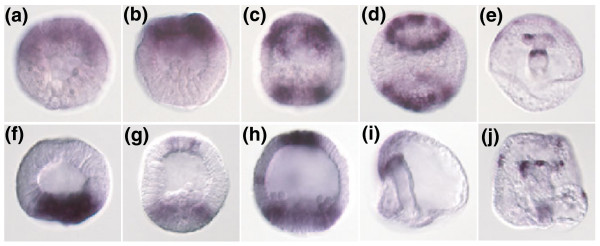
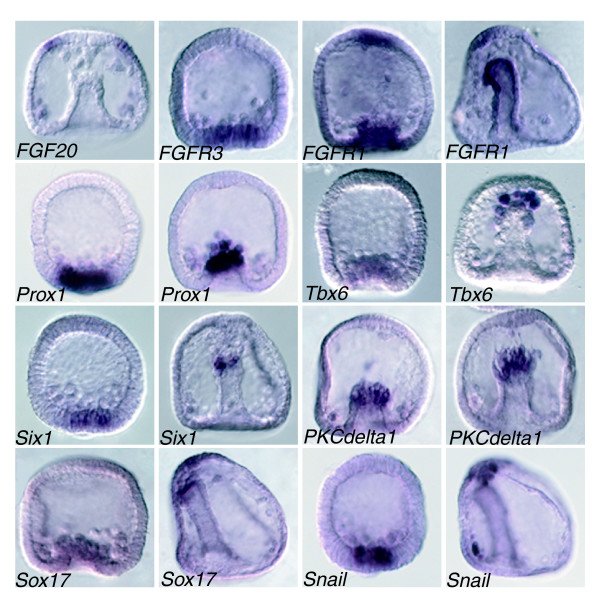
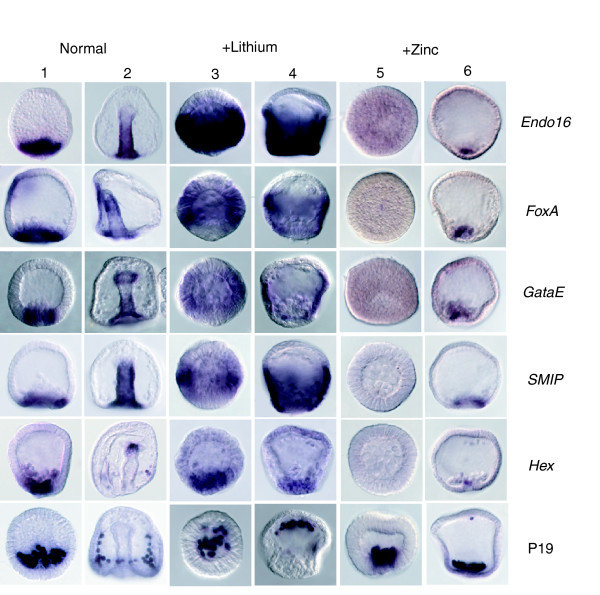
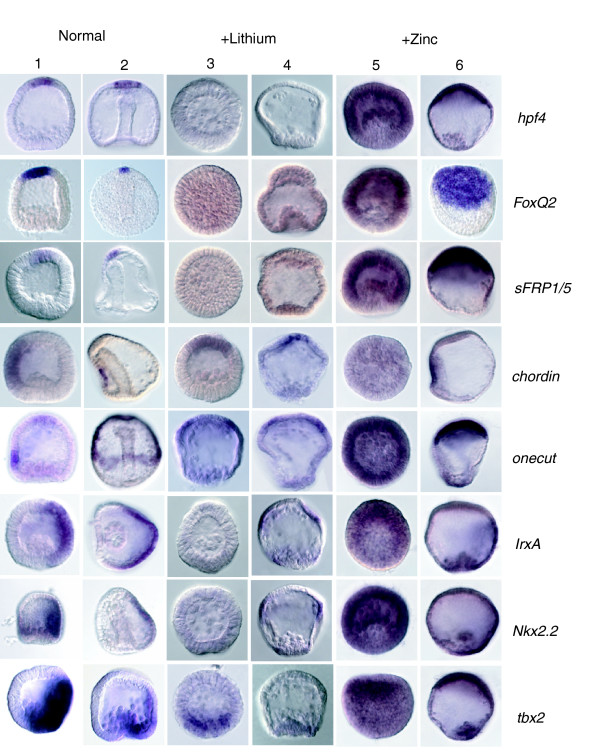
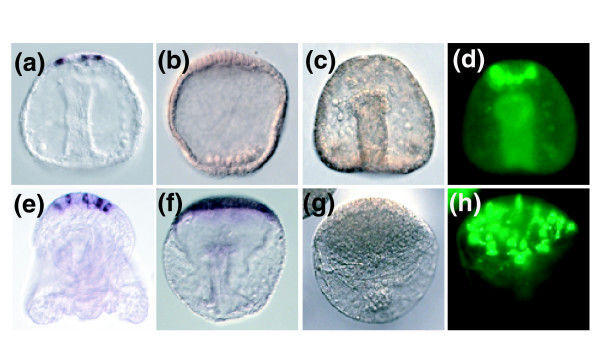
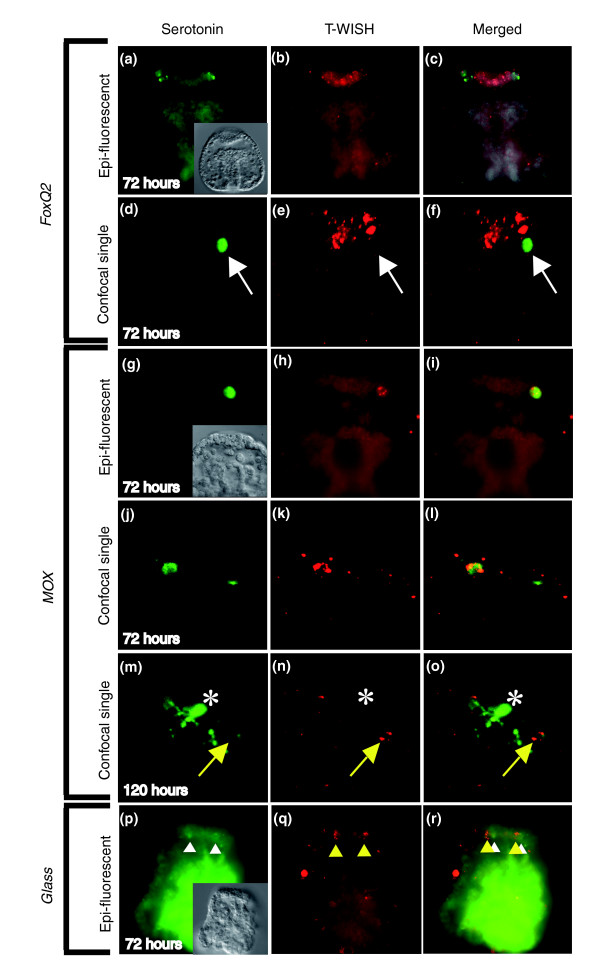
Results: Using large-scale screens on arrays carrying 50% to 70% of all genes, we identified novel territory-specific markers. Our strategy was based on computational selection of genes that are differentially expressed in lithium-treated embryos, which form excess endomesoderm, and in zinc-treated embryos, in which endomesoderm specification is blocked. Whole-mount in situ hybridization (WISH) analysis of 700 genes indicates that the apical organ region is eliminated in lithium-treated embryos. Conversely, apical and specifically neural markers are expressed more broadly in zinc-treated embryos, whereas endomesoderm signaling is severely reduced. Strikingly, the number of serotonergic neurons is amplified by at least tenfold in zinc-treated embryos. WISH analysis further indicates that there is crosstalk between the Wnt (wingless int), Notch, and fibroblast growth factor signaling pathways in secondary mesoderm cell specification and differentiation, similar to signaling cascades that function during development of presomitic mesoderm in mouse embryogenesis. We provide differential expression data for more than 4,000 genes and WISH patterns of more than 250 genes, and more than 2,400 annotated WISH images.
Conclusion: Our work provides tissue-specific expression patterns for a large fraction of the sea urchin genes that have not yet been included in existing regulatory networks and await functional integration. Furthermore, we noted neuron-inducing activity of zinc on embryonic development; this is the first observation of such activity in any organism.
Figures
Similar articles
-
New early zygotic regulators expressed in endomesoderm of sea urchin embryos discovered by differential array hybridization.Dev Biol. 2002 Jun 1;246(1):132-47. doi: 10.1006/dbio.2002.0607. Dev Biol. 2002. PMID: 12027439
-
Signal-dependent regulation of the sea urchin skeletogenic gene regulatory network.Gene Expr Patterns. 2014 Nov;16(2):93-103. doi: 10.1016/j.gep.2014.10.002. Epub 2014 Oct 16. Gene Expr Patterns. 2014. PMID: 25460514
-
Specific functions of the Wnt signaling system in gene regulatory networks throughout the early sea urchin embryo.Proc Natl Acad Sci U S A. 2014 Nov 25;111(47):E5029-38. doi: 10.1073/pnas.1419141111. Epub 2014 Nov 10. Proc Natl Acad Sci U S A. 2014. PMID: 25385617 Free PMC article.
-
Gene regulatory networks for ectoderm specification in sea urchin embryos.Biochim Biophys Acta. 2009 Apr;1789(4):261-7. doi: 10.1016/j.bbagrm.2009.02.002. Epub 2009 Mar 21. Biochim Biophys Acta. 2009. PMID: 19429544 Review.
-
Gene regulation: gene control network in development.Annu Rev Biophys Biomol Struct. 2007;36:191. doi: 10.1146/annurev.biophys.35.040405.102002. Annu Rev Biophys Biomol Struct. 2007. PMID: 17291181 Review.
Cited by
-
Notch and Nodal control forkhead factor expression in the specification of multipotent progenitors in sea urchin.Development. 2013 Apr;140(8):1796-806. doi: 10.1242/dev.091157. Development. 2013. PMID: 23533178 Free PMC article.
-
Evolutionarily conserved Wnt/Sp5 signaling is critical for anterior-posterior axis patterning in sea urchin embryos.iScience. 2023 Dec 2;27(1):108616. doi: 10.1016/j.isci.2023.108616. eCollection 2024 Jan 19. iScience. 2023. PMID: 38179064 Free PMC article.
-
The bilaterian head patterning gene six3/6 controls aboral domain development in a cnidarian.PLoS Biol. 2013;11(2):e1001488. doi: 10.1371/journal.pbio.1001488. Epub 2013 Feb 19. PLoS Biol. 2013. PMID: 23483856 Free PMC article.
-
A single cell RNA sequencing resource for early sea urchin development.Development. 2020 Sep 11;147(17):dev191528. doi: 10.1242/dev.191528. Development. 2020. PMID: 32816969 Free PMC article.
-
Spatial and temporal patterns of gene expression during neurogenesis in the sea urchin Lytechinus variegatus.Evodevo. 2019 Feb 12;10:2. doi: 10.1186/s13227-019-0115-8. eCollection 2019. Evodevo. 2019. PMID: 30792836 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
