

MyD88-dependent immune activation mediated by human immunodeficiency virus type 1-encoded Toll-like receptor ligands
- PMID: 17507480
- PMCID: PMC1951290
- DOI: 10.1128/JVI.00421-07
MyD88-dependent immune activation mediated by human immunodeficiency virus type 1-encoded Toll-like receptor ligands
Abstract
Immune activation is a major characteristic of human immunodeficiency virus type 1 (HIV-1) infection and a strong prognostic factor for HIV-1 disease progression. The underlying mechanisms leading to immune activation in viremic HIV-1 infection, however, are not fully understood. Here we show that, following the initiation of highly active antiretroviral therapy, the immediate decline of immune activation is closely associated with the reduction of HIV-1 viremia, which suggests a direct contribution of HIV-1 itself to immune activation. To propose a mechanism, we demonstrate that the single-stranded RNA of HIV-1 encodes multiple uridine-rich Toll-like receptor 7/8 (TLR7/8) ligands that induce strong MyD88-dependent plasmacytoid dendritic cell and monocyte activation, as well as accessory cell-dependent T-cell activation. HIV-1-encoded TLR ligands may, therefore, directly contribute to the immune activation observed during viremic HIV-1 infection. These data provide an initial rationale for inhibiting the TLR pathway to directly reduce the chronic immune activation induced by HIV-1 and the associated immune pathogenesis.
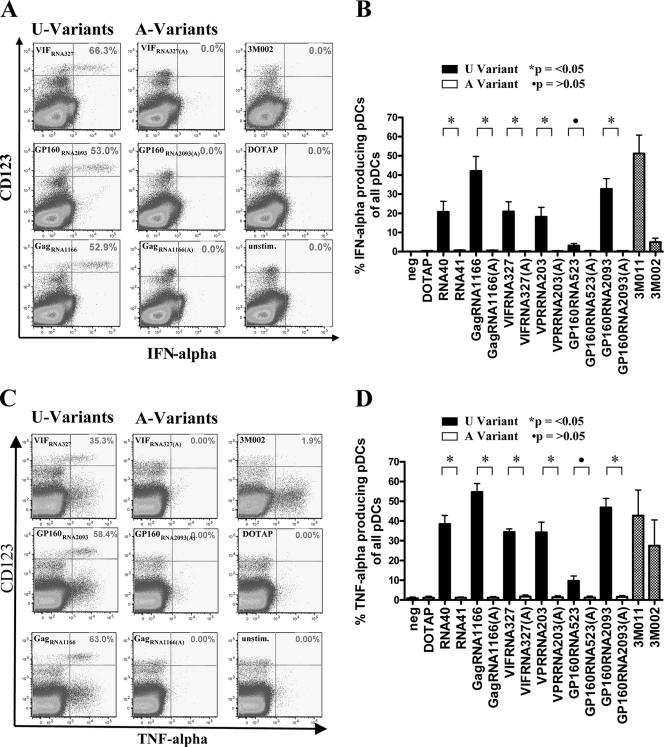
Figures







Similar articles
-
Retrovirus-Based Virus-Like Particle Immunogenicity and Its Modulation by Toll-Like Receptor Activation.J Virol. 2017 Oct 13;91(21):e01230-17. doi: 10.1128/JVI.01230-17. Print 2017 Nov 1. J Virol. 2017. PMID: 28794025 Free PMC article.
-
Single-stranded RNA derived from HIV-1 serves as a potent activator of NK cells.J Immunol. 2007 Jun 15;178(12):7658-66. doi: 10.4049/jimmunol.178.12.7658. J Immunol. 2007. PMID: 17548602
-
Increased expression of TLR7 in CD8(+) T cells leads to TLR7-mediated activation and accessory cell-dependent IFN-gamma production in HIV type 1 infection.AIDS Res Hum Retroviruses. 2009 Dec;25(12):1287-95. doi: 10.1089/aid.2008.0303. AIDS Res Hum Retroviruses. 2009. PMID: 19954299
-
Molecular biological assessment methods and understanding the course of the HIV infection.APMIS Suppl. 2003;(114):1-37. APMIS Suppl. 2003. PMID: 14626050 Review.
-
Immune activation and the role of TLRs and TLR agonists in the pathogenesis of HIV-1 infection in the humanized mouse model.J Infect Dis. 2013 Nov;208 Suppl 2(Suppl 2):S145-9. doi: 10.1093/infdis/jit402. J Infect Dis. 2013. PMID: 24151321 Free PMC article. Review.
Cited by
-
Higher expression of several interferon-stimulated genes in HIV-1-infected females after adjusting for the level of viral replication.J Infect Dis. 2013 Sep 1;208(5):830-8. doi: 10.1093/infdis/jit262. Epub 2013 Jun 10. J Infect Dis. 2013. PMID: 23757341 Free PMC article.
-
Interplay between Intrinsic and Innate Immunity during HIV Infection.Cells. 2019 Aug 17;8(8):922. doi: 10.3390/cells8080922. Cells. 2019. PMID: 31426525 Free PMC article. Review.
-
HIV-1 exploits innate signaling by TLR8 and DC-SIGN for productive infection of dendritic cells.Nat Immunol. 2010 May;11(5):419-26. doi: 10.1038/ni.1858. Epub 2010 Apr 4. Nat Immunol. 2010. PMID: 20364151
-
RIG-I-mediated antiviral signaling is inhibited in HIV-1 infection by a protease-mediated sequestration of RIG-I.J Virol. 2011 Feb;85(3):1224-36. doi: 10.1128/JVI.01635-10. Epub 2010 Nov 17. J Virol. 2011. PMID: 21084468 Free PMC article.
-
HIV-1 activates macrophages independent of Toll-like receptors.PLoS One. 2008;3(12):e3664. doi: 10.1371/journal.pone.0003664. Epub 2008 Dec 2. PLoS One. 2008. PMID: 19048100 Free PMC article.
References
-
- Akira, S., K. Takeda, and T. Kaisho. 2001. Toll-like receptors: critical proteins linking innate and acquired immunity. Nat. Immunol. 2:675-680. - PubMed
-
- Beignon, A. S., K. McKenna, M. Skoberne, O. Manches, I. Dasilva, D. G. Kavanagh, M. Larsson, R. J. Gorelick, J. D. Lifson, and N. Bhardwaj. 2005. Endocytosis of HIV-1 activates plasmacytoid dendritic cells via Toll-like receptor-viral RNA interactions. J. Clin. Investig. 115:3265-3275. - PMC - PubMed
-
- Bekeredjian-Ding, I., S. I. Roth, S. Gilles, T. Giese, A. Ablasser, V. Hornung, S. Endres, and G. Hartmann. 2006. T cell-independent, TLR-induced IL-12p70 production in primary human monocytes. J. Immunol. 176:7438-7446. - PubMed
-
- Bisset, L. R., R. W. Cone, W. Huber, M. Battegay, P. L. Vernazza, R. Weber, P. J. Grob, and M. Opravil. 1998. Highly active antiretroviral therapy during early HIV infection reverses T-cell activation and maturation abnormalities. Swiss HIV Cohort Study. AIDS 12:2115-2123. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
