

Thermolabilizing pseudoreversions in reovirus outer-capsid protein micro 1 rescue the entry defect conferred by a thermostabilizing mutation
- PMID: 17507494
- PMCID: PMC1933377
- DOI: 10.1128/JVI.02720-06
Thermolabilizing pseudoreversions in reovirus outer-capsid protein micro 1 rescue the entry defect conferred by a thermostabilizing mutation
Abstract
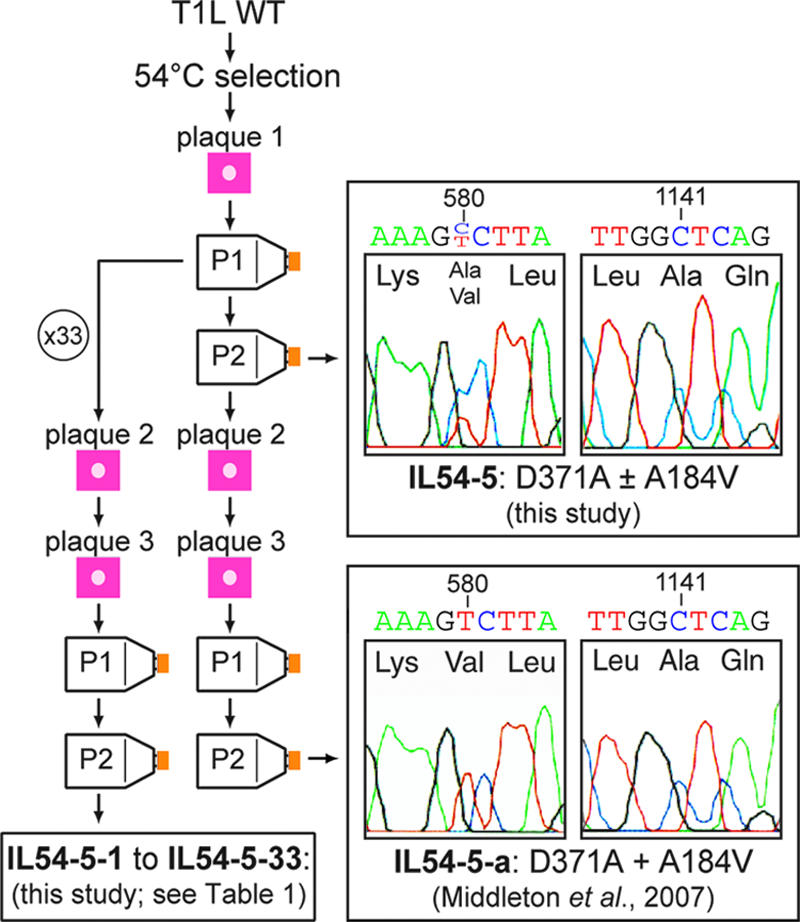
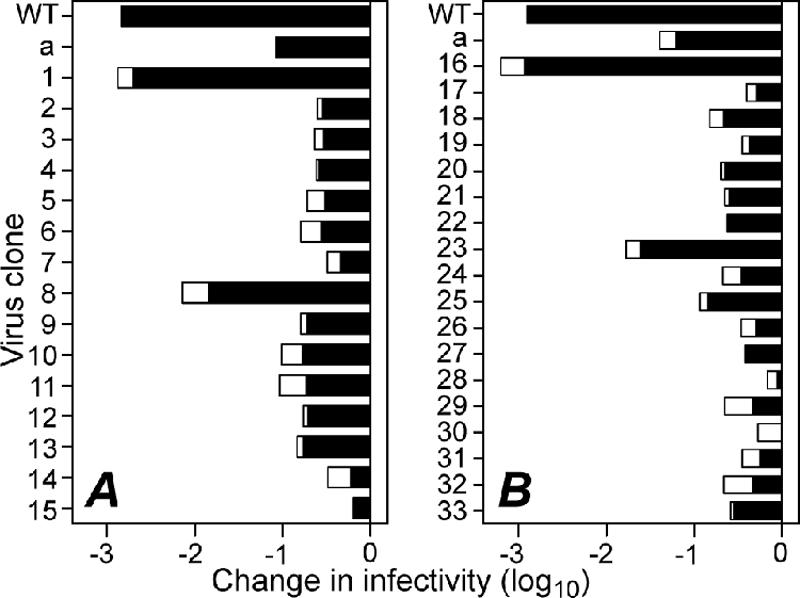
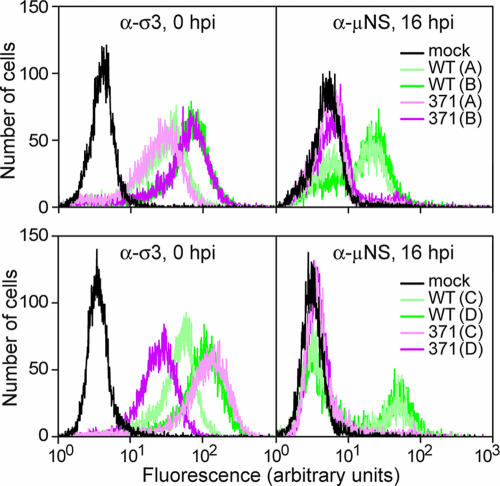
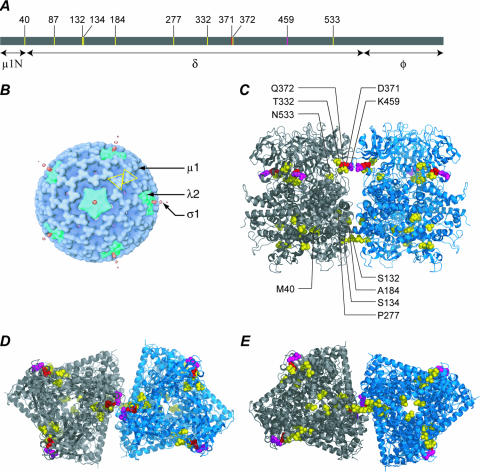
Heat-resistant mutants selected from infectious subvirion particles of mammalian reoviruses have determinative mutations in the major outer-capsid protein micro 1. Here we report the isolation and characterization of intragenic pseudoreversions of one such thermostabilizing mutation. From a plaque that had survived heat selection, a number of viruses with one shared mutation but different second-site mutations were isolated. The effect of the shared mutation alone or in combination with second-site mutations was examined using recoating genetics. The shared mutation, D371A, was found to confer (i) substantial thermostability, (ii) an infectivity defect that followed attachment but preceded viral protein synthesis, and (iii) resistance to micro 1 rearrangement in vitro, with an associated failure to lyse red blood cells. Three different second-site mutations were individually tested in combination with D371A and found to wholly or partially revert these phenotypes. Furthermore, when tested alone in recoated particles, each of these three second-site mutations conferred demonstrable thermolability. This and other evidence suggest that pseudoreversion of micro 1-based thermostabilization can occur by a general mechanism of micro 1-based thermolabilization, not requiring a specific compensatory mutation. The thermostabilizing mutation D371A as well as 9 of the 10 identified second-site mutations are located near contact regions between micro 1 trimers in the reovirus outer capsid. The availability of both thermostabilizing and thermolabilizing mutations in micro 1 should aid in defining the conformational rearrangements and mechanisms involved in membrane penetration during cell entry by this structurally complex nonenveloped animal virus.
Figures
References
-
- Borsa, J., D. G. Long, T. P. Copps, M. D. Sargent, and J. D. Chapman. 1974. Reovirus transcriptase activation in vitro: further studies on the facilitation phenomenon. Intervirology 3:15-35. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
