

Staufen1 regulates diverse classes of mammalian transcripts
- PMID: 17510634
- PMCID: PMC1888674
- DOI: 10.1038/sj.emboj.7601712
Staufen1 regulates diverse classes of mammalian transcripts
Abstract
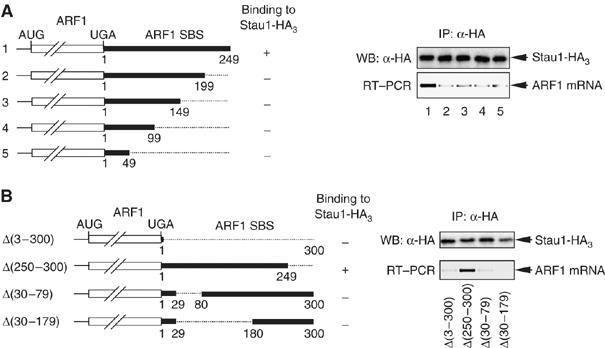
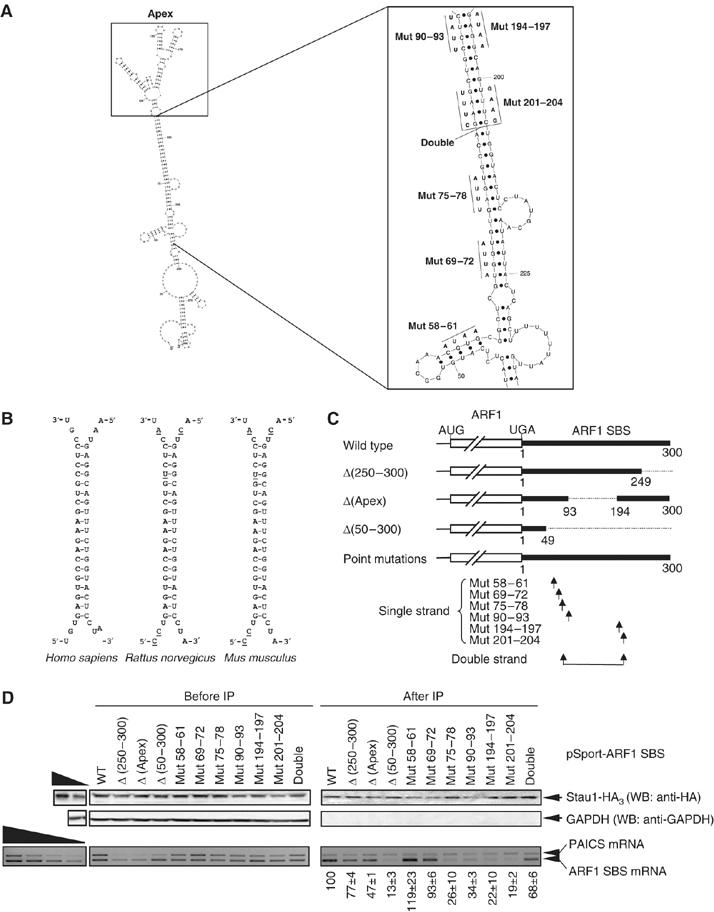
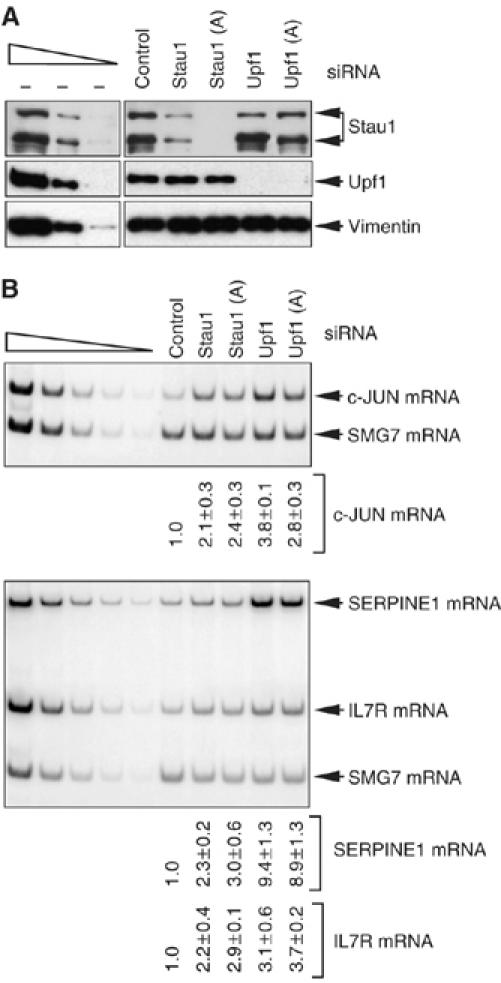
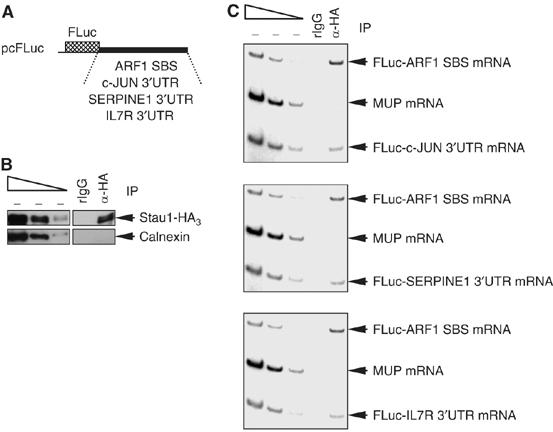
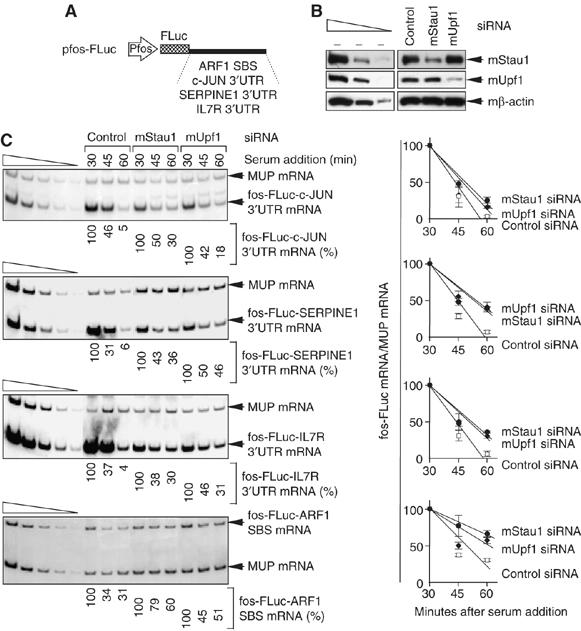
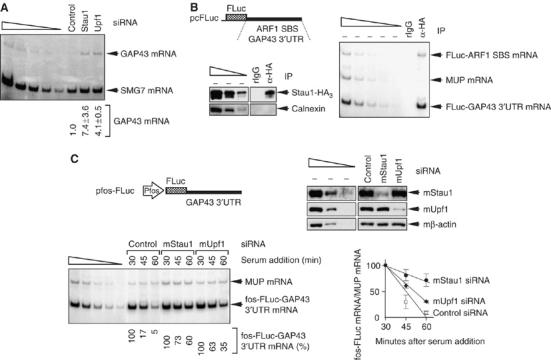
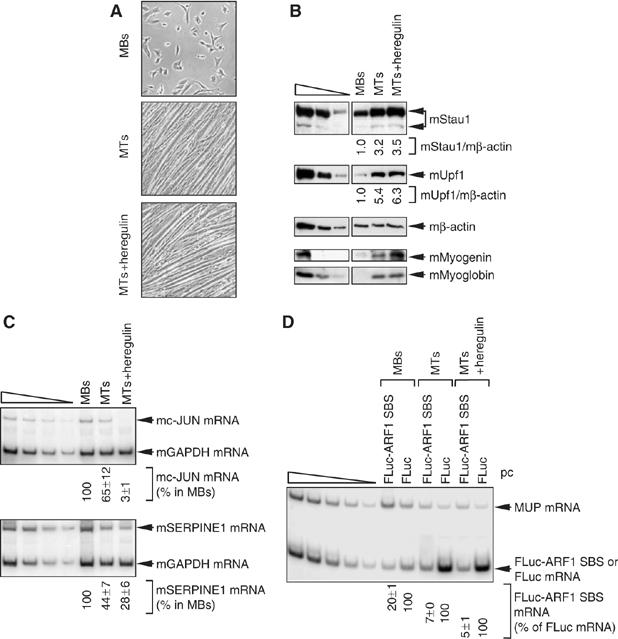
It is currently unknown how extensively the double-stranded RNA-binding protein Staufen (Stau)1 is utilized by mammalian cells to regulate gene expression. To date, Stau1 binding to the 3'-untranslated region (3'-UTR) of ADP ribosylation factor (ARF)1 mRNA has been shown to target ARF1 mRNA for Stau1-mediated mRNA decay (SMD). ARF1 SMD depends on translation and recruitment of the nonsense-mediated mRNA decay factor Upf1 to the ARF1 3'-UTR by Stau1. Here, we demonstrate that Stau1 binds to a complex structure within the ARF1 3'-UTR. We also use microarrays to show that 1.1 and 1.0% of the 11 569 HeLa-cell transcripts that were analyzed are upregulated and downregulated, respectively, at least two-fold upon Stau1 depletion in three independently performed experiments. We localize the Stau1 binding site to the 3'-UTR of four mRNAs that we define as natural SMD targets. Additionally, we provide evidence that the efficiency of SMD increases during the differentiation of C2C12 myoblasts to myotubes. We propose that Stau1 influences the expression of a wide variety of physiologic transcripts and metabolic pathways.
Figures
References
-
- Bélanger G, Stocksley MA, Vandromme M, Schaeffer L, Furic L, DesGroseillers L, Jasmin BJ (2003) Localization of the RNA-binding proteins Staufen1 and Staufen2 at the mammalian neuromuscular junction. J Neurochem 86: 669–677 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
