

Disruption of erythroid K-Cl cotransporters alters erythrocyte volume and partially rescues erythrocyte dehydration in SAD mice
- PMID: 17510708
- PMCID: PMC1866252
- DOI: 10.1172/JCI30630
Disruption of erythroid K-Cl cotransporters alters erythrocyte volume and partially rescues erythrocyte dehydration in SAD mice
Abstract
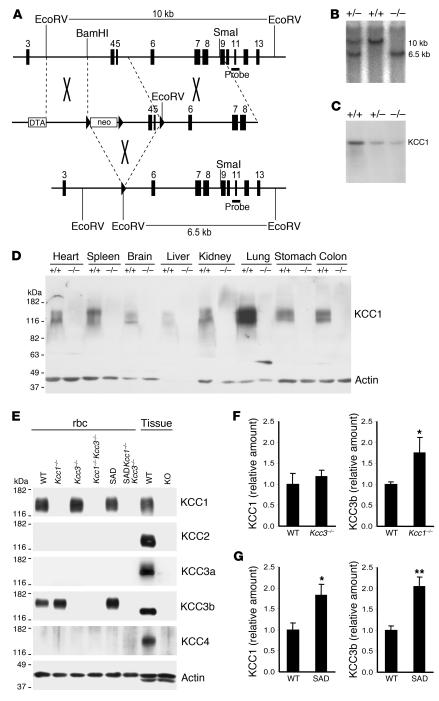
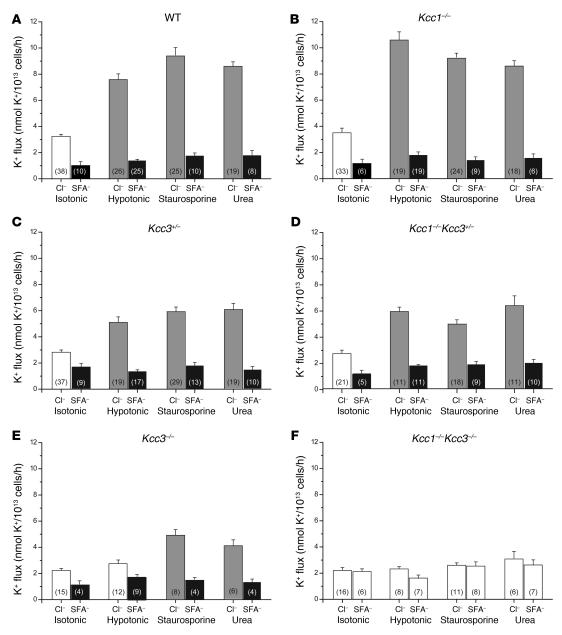
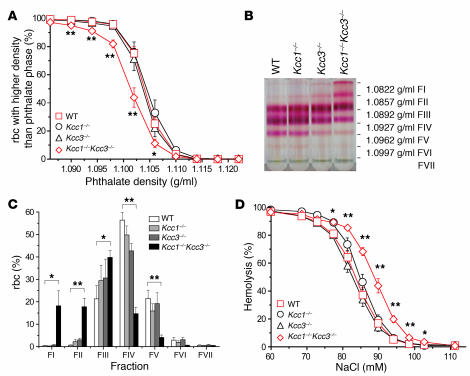
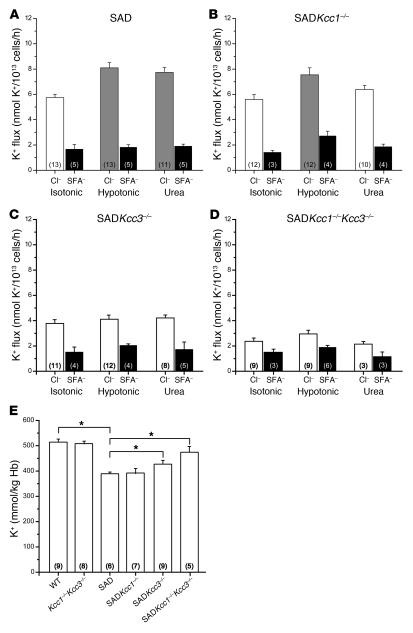
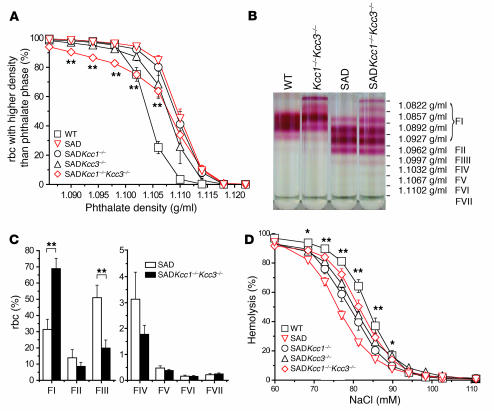
K-Cl cotransport activity in rbc is a major determinant of rbc volume and density. Pathologic activation of erythroid K-Cl cotransport activity in sickle cell disease contributes to rbc dehydration and cell sickling. To address the roles of individual K-Cl cotransporter isoforms in rbc volume homeostasis, we disrupted the Kcc1 and Kcc3 genes in mice. As rbc K-Cl cotransport activity was undiminished in Kcc1(-/-) mice, decreased in Kcc3(-/-) mice, and almost completely abolished in mice lacking both isoforms, we conclude that K-Cl cotransport activity of mouse rbc is mediated largely by KCC3. Whereas rbc of either Kcc1(-/-) or Kcc3(-/-) mice were of normal density, rbc of Kcc1(-/-)Kcc3(-/-) mice exhibited defective volume regulation, including increased mean corpuscular volume, decreased density, and increased susceptibility to osmotic lysis. K-Cl cotransport activity was increased in rbc of SAD mice, which are transgenic for a hypersickling human hemoglobin S variant. Kcc1(-/-)Kcc3(-/-) SAD rbc lacked nearly all K-Cl cotransport activity and exhibited normalized values of mean corpuscular volume, corpuscular hemoglobin concentration mean, and K(+) content. Although disruption of K-Cl cotransport rescued the dehydration phenotype of most SAD rbc, the proportion of the densest red blood cell population remained unaffected.
Figures
References
-
- Lauf P.K., Theg B.E. A chloride dependent K+ flux induced by N-ethylmaleimide in genetically low K+ sheep and goat erythrocytes. . Biochem. Biophys. Res. Commun. 1980;92:1422–1428. - PubMed
-
- Gamba G. Molecular physiology and pathophysiology of electroneutral cation-chloride cotransporters. Physiol. Rev. 2005;85:423–493. - PubMed
-
- Howard H.C., et al. The K-Cl cotransporter KCC3 is mutant in a severe peripheral neuropathy associated with agenesis of the corpus callosum. Nat. Genet. 2002;32:384–392. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
