

The Sec14-superfamily and the regulatory interface between phospholipid metabolism and membrane trafficking
- PMID: 17512778
- PMCID: PMC2001170
- DOI: 10.1016/j.bbalip.2007.04.002
The Sec14-superfamily and the regulatory interface between phospholipid metabolism and membrane trafficking
Abstract
A central principle of signal transduction is the appropriate control of the process so that relevant signals can be detected with fine spatial and temporal resolution. In the case of lipid-mediated signaling, organization and metabolism of specific lipid mediators is an important aspect of such control. Herein, we review the emerging evidence regarding the roles of Sec14-like phosphatidylinositol transfer proteins (PITPs) in the action of intracellular signaling networks; particularly as these relate to membrane trafficking. Finally, we explore developing ideas regarding how Sec14-like PITPs execute biological function. As Sec14-like proteins define a protein superfamily with diverse lipid (or lipophile) binding capabilities, it is likely these under-investigated proteins will be ultimately demonstrated as a ubiquitously important set of biological regulators whose functions influence a large territory in the signaling landscape of eukaryotic cells.
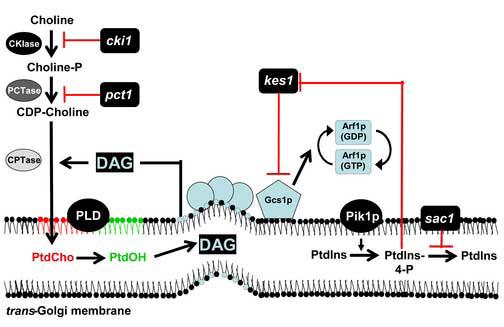
Figures




References
-
- Bankaitis VA, Aitken JR, Cleves AE, Dowhan W. An essential role for a phospholipid transfer protein in yeast Golgi function. Nature. 1990;347(6293):561–2. - PubMed
-
- Phillips SE, Vincent P, Rizzieri KE, Schaaf G, Gaucher EA, Bankaitis VA. The diverse biological functions of phosphatidylinositol transfer proteins in eukaryotes. Crit Rev Biochem Mol Biol. 2005;41(1):21–49. - PubMed
-
- Novick P, Field C, Schekman R. Identification of 23 complementation groups required for post-translational events in the yeast secretory pathway. Cell. 1980;21(1):205–15. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
