

Components of the Arabidopsis mRNA decapping complex are required for early seedling development
- PMID: 17513503
- PMCID: PMC1913740
- DOI: 10.1105/tpc.106.047621
Components of the Arabidopsis mRNA decapping complex are required for early seedling development
Abstract
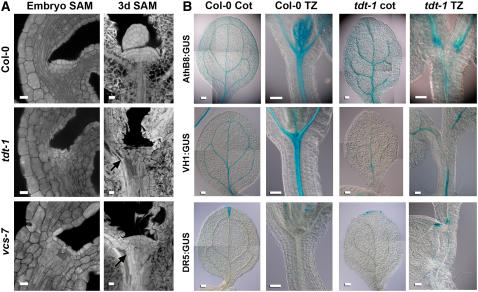
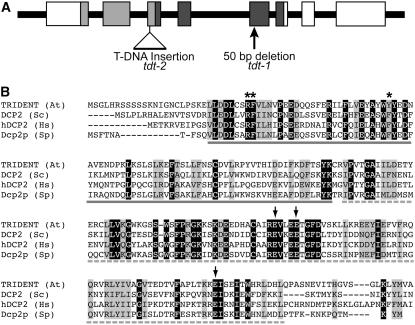
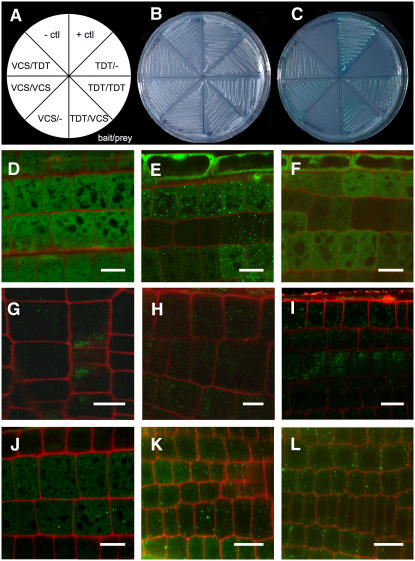
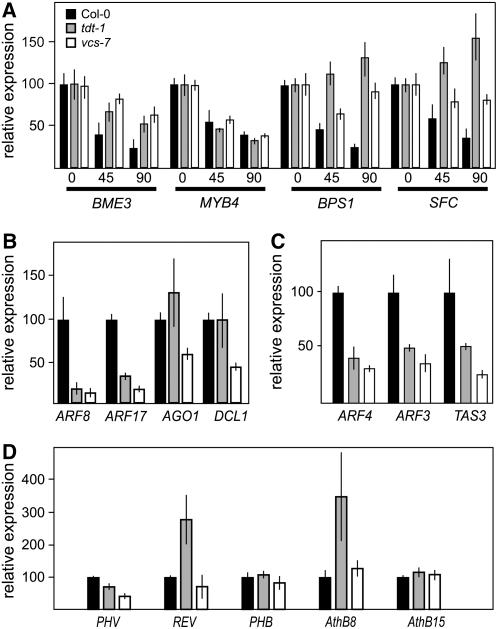
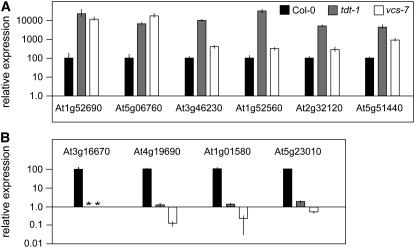
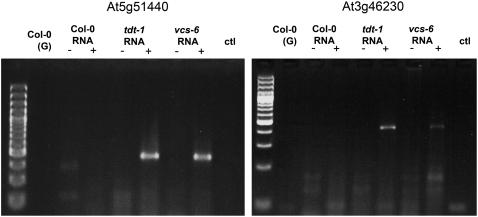
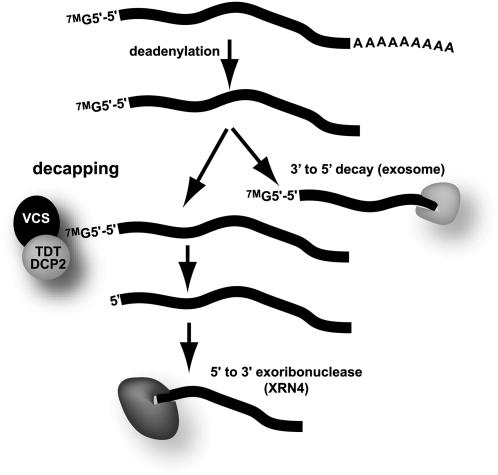
To understand the mechanisms controlling vein patterning in Arabidopsis thaliana, we analyzed two phenotypically similar mutants, varicose (vcs) and trident (tdt). We had previously identified VCS, and recently, human VCS was shown to function in mRNA decapping. Here, we report that TDT encodes the mRNA-decapping enzyme. VCS and TDT function together in small cytoplasmic foci that appear to be processing bodies. To understand the developmental requirements for mRNA decapping, we characterized the vcs and tdt phenotypes. These mutants were small and chlorotic, with severe defects in shoot apical meristem formation and cotyledon vein patterning. Many capped mRNAs accumulated in tdt and vcs mutants, but surprisingly, some mRNAs were specifically depleted. In addition, loss of decapping arrested the decay of some mRNAs, while others showed either modest or no decay defects, suggesting that mRNAs may show specificity for particular decay pathways (3' to 5' and 5' to 3'). Furthermore, the severe block to postembryonic development in vcs and tdt and the accompanying accumulation of embryonic mRNAs indicate that decapping is important for the embryo-to-seedling developmental transition.
Figures

References
-
- Adenot, X., Elmayan, T., Lauressergues, D., Boutet, S., Bouché, N., Gasciolli, V., and Vaucheret, H. (2006). DRB4-dependent TAS3 trans-acting siRNAs control leaf morphology through AGO7. Curr. Biol. 16 927–932. - PubMed
-
- Alonso, J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301 653–657. - PubMed
-
- Badis, G., Saveanu, C., Fromont-Racine, M., and Jacquier, A. (2004). Targeted mRNA degradation by deadenylation-independent decapping. Mol. Cell 15 5–15. - PubMed
-
- Baima, S., Nobili, F., Sessa, G., Lucchetti, S., Ruberti, I., and Morelli, G. (1995). The expression of the Athb-8 homeobox gene is restricted to provascular cells in Arabidopsis thaliana. Development 121 4171–4182. - PubMed
-
- Barton, M.K., and Poethig, R.S. (1993). Formation of the shoot apical meristem in Arabidopsis thaliana: An analysis of development in the wild type and in the shoot meristemless mutant. Development 119 823–831.
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
