

A novel splicing factor, Yju2, is associated with NTC and acts after Prp2 in promoting the first catalytic reaction of pre-mRNA splicing
- PMID: 17515604
- PMCID: PMC1952081
- DOI: 10.1128/MCB.00346-07
A novel splicing factor, Yju2, is associated with NTC and acts after Prp2 in promoting the first catalytic reaction of pre-mRNA splicing
Abstract
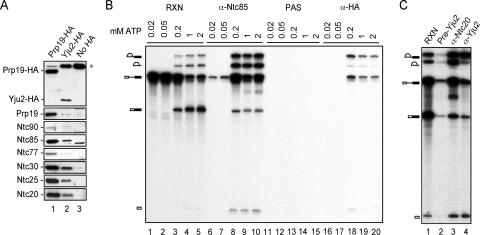
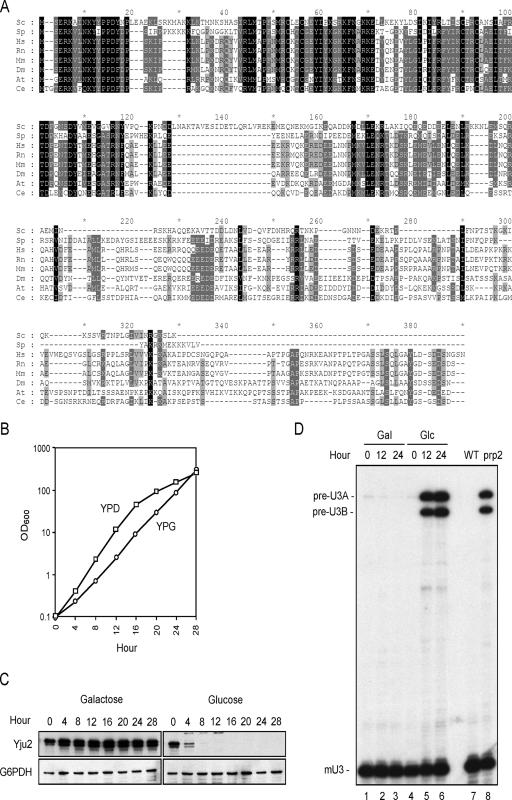
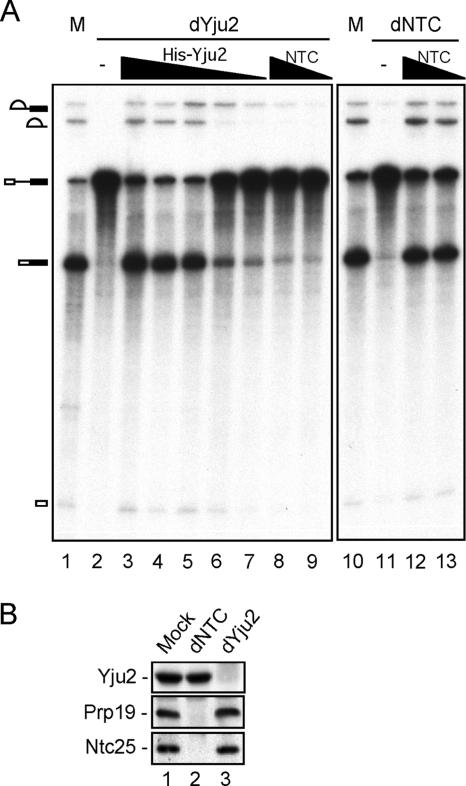
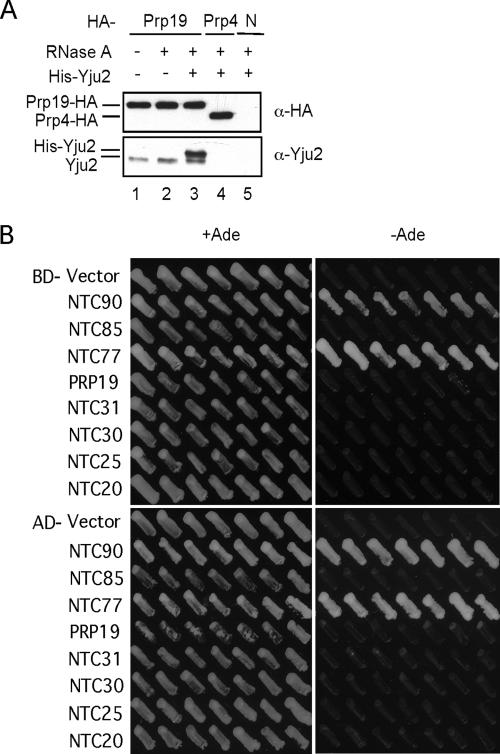
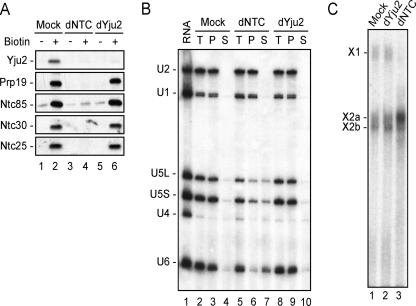
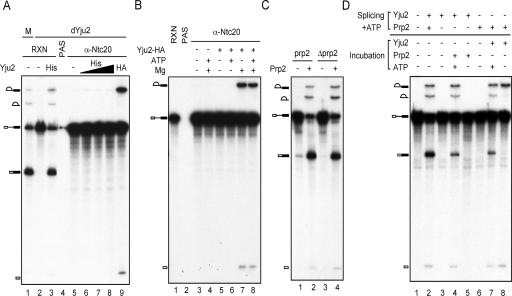
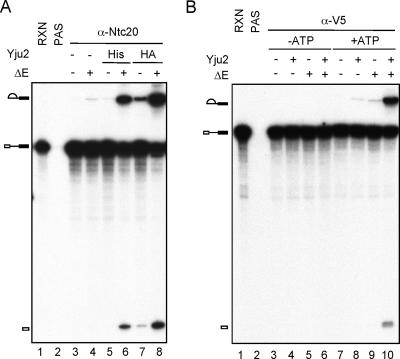
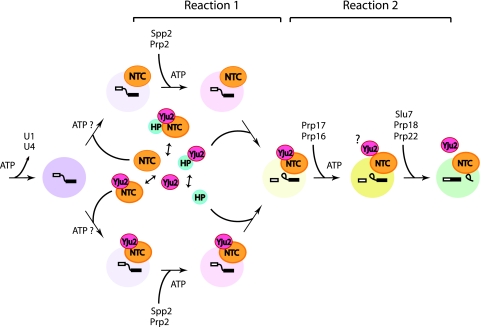
The Prp19-associated complex (NTC) is essential for pre-mRNA splicing and is associated with the spliceosome during spliceosome activation. NTC is required for specifying interactions of U5 and U6 with pre-mRNA to stabilize their association with the spliceosome after dissociation of U4. Here, we show that a novel splicing factor, Yju2, is associated with components of NTC, and that it is required for pre-mRNA splicing both in vivo and in vitro. During spliceosome assembly, Yju2 is associated with the spliceosome at nearly the same time as NTC but is destabilized after the first catalytic reaction, whereas other NTC components remain associated until the reaction is complete. Extracts depleted of Yju2 could be complemented by recombinant Yju2, suggesting that Yju2 and NTC are not entirely in association with each other. Yju2 is not required for the binding of NTC to the spliceosome or for NTC-mediated spliceosome activation. Complementation analysis of the affinity-isolated spliceosome formed in Yju2-depleted extracts demonstrated that Yju2 acts in concert with an unidentified heat-resistant factor(s) in an ATP-independent manner to promote the first catalytic reaction of pre-mRNA splicing after Prp2-mediated structural rearrangement of the spliceosome.
Figures
References
-
- Brow, D. A. 2002. Allosteric cascade of spliceosome activation. Annu. Rev. Genet. 36:333-360. - PubMed
-
- Burge, C. B., T. H. Tuschl, and P. A. Sharp. 1999. Splicing of precursors to mRNAs by the spliceosome, p. 525-560. In R. F. Gesteland, T. R. Cech, and J. F. Atkins (ed.), RNA world II. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
-
- Chan, S.-P., and S.-C. Cheng. 2005. The Prp19-associated complex is required for specifying interactions of U5 and U6 with pre-mRNA during spliceosome activation. J. Biol. Chem. 280:31190-31199. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous