

Pph3-Psy2 is a phosphatase complex required for Rad53 dephosphorylation and replication fork restart during recovery from DNA damage
- PMID: 17517611
- PMCID: PMC1890487
- DOI: 10.1073/pnas.0703252104
Pph3-Psy2 is a phosphatase complex required for Rad53 dephosphorylation and replication fork restart during recovery from DNA damage
Abstract
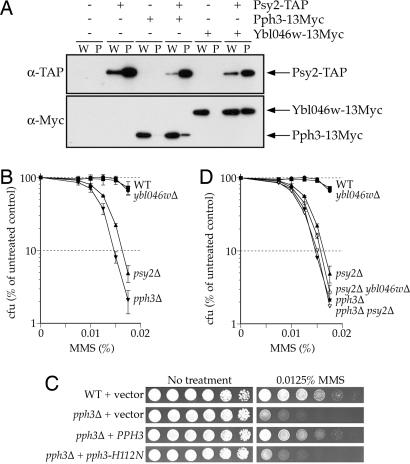
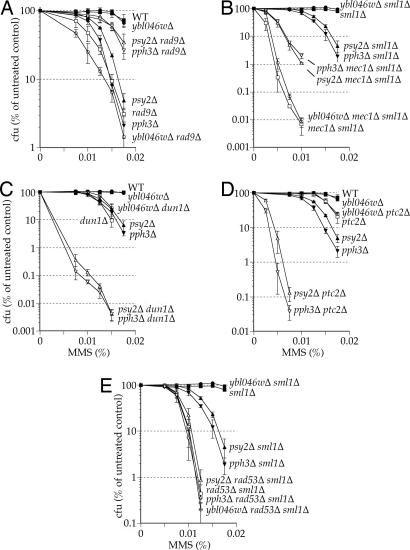
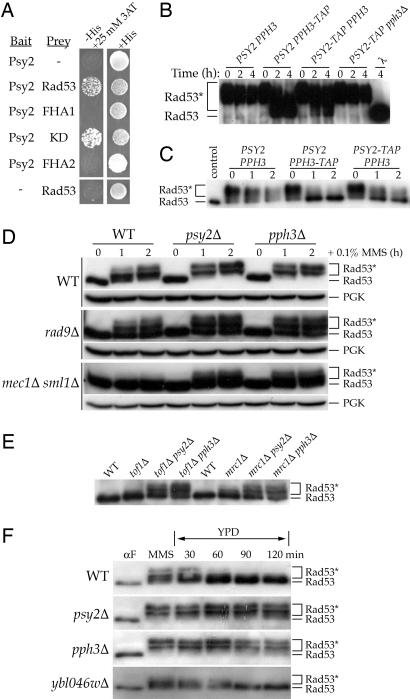
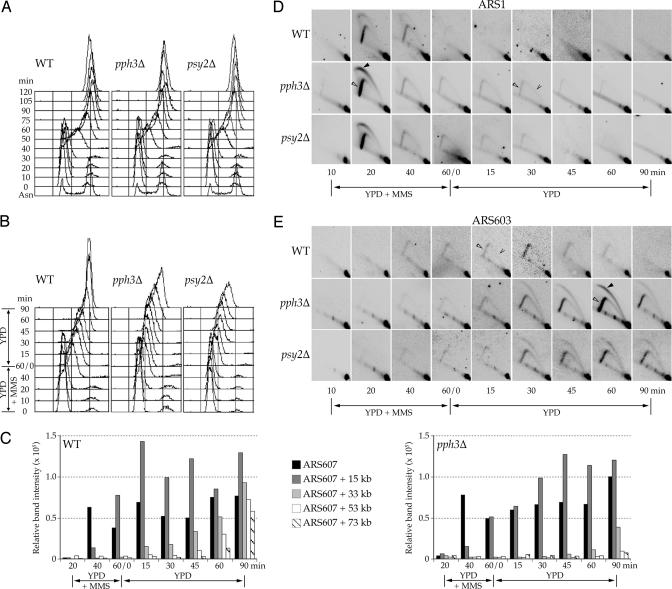
Activation of the checkpoint kinase Rad53 is a critical response to DNA damage that results in stabilization of stalled replication forks, inhibition of late-origin initiation, up-regulation of dNTP levels, and delayed entry to mitosis. Activation of Rad53 is well understood and involves phosphorylation by the protein kinases Mec1 and Tel1 as well as in trans autophosphorylation by Rad53 itself. However, deactivation of Rad53, which must occur to allow the cell to recover from checkpoint arrest, is not well understood. Here, we present genetic and biochemical evidence that the type 2A-like protein phosphatase Pph3 forms a complex with Psy2 (Pph3-Psy2) that binds and dephosphorylates activated Rad53 during treatment with, and recovery from, methylmethane sulfonate-mediated DNA damage. In the absence of Pph3-Psy2, Rad53 dephosphorylation and the resumption of DNA synthesis are delayed during recovery from DNA damage. This delay in DNA synthesis reflects a failure to restart stalled replication forks, whereas, remarkably, genome replication is eventually completed by initiating late origins of replication despite the presence of hyperphosphorylated Rad53. These findings suggest that Rad53 regulates replication fork restart and initiation of late firing origins independently and that regulation of these processes is mediated by specific Rad53 phosphatases.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Kolodner RD, Putnam CD, Myung K. Science. 2002;297:552–557. - PubMed
-
- Nyberg KA, Michelson RJ, Putnam CW, Weinert TA. Annu Rev Genet. 2002;36:617–656. - PubMed
-
- Lambert S, Carr AM. Biochimie. 2005;87:591–602. - PubMed
-
- Branzei D, Foiani M. Curr Opin Cell Biol. 2005;17:568–575. - PubMed
-
- Ma J-L, Lee S-J, Duong JK, Stern DF. J Biol Chem. 2006;281:3954–3963. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
