

Roles of heat-shock protein 90 in maintaining and facilitating the neurodegenerative phenotype in tauopathies
- PMID: 17517623
- PMCID: PMC1890525
- DOI: 10.1073/pnas.0701055104
Roles of heat-shock protein 90 in maintaining and facilitating the neurodegenerative phenotype in tauopathies
Abstract
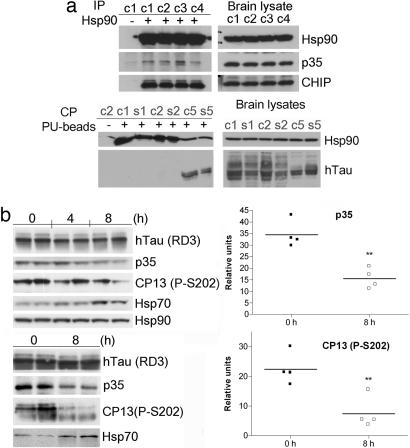
Neurodegeneration, a result of multiple dysregulatory events, is a lengthy multistep process manifested by accrual of mutant variants and abnormal expression, posttranslational modification, and processing of certain proteins. Accumulation of these dysregulated processes requires a mechanism that maintains their functional stability and allows the evolution of the neurodegenerative phenotype. In malignant cells, the capacity to buffer transformation has been attributed to heat-shock protein 90 (Hsp90). Although normal proteins seem to require limited assistance from the chaperone, their aberrant counterparts seem to be highly dependent on Hsp90. Whereas enhanced Hsp90 affinity for mutated or functionally deregulated client proteins has been observed for several oncoproteins, it is unknown whether Hsp90 plays a similar role for neuronal proteins and thus maintains and facilitates the transformed phenotype in neurodegenerative diseases. Tauopathies are neurodegenerative diseases characterized by aberrant phosphorylation and/or expression of Tau protein, leading to a time-dependent accumulation of Tau aggregates and subsequent neuronal death. Here, we show that the stability of p35, a neuronal protein that activates cyclin-dependent protein kinase 5 through complex formation leading to aberrant Tau phosphorylation, and that of mutant but not WT Tau protein is maintained in tauopathies by Hsp90. Inhibition of Hsp90 in cellular and mouse models of tauopathies leads to a reduction of the pathogenic activity of these proteins and results in elimination of aggregated Tau. The results identify important roles played by Hsp90 in maintaining and facilitating the degenerative phenotype in these diseases and provide a common principle governing cancer and neurodegenerative diseases.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
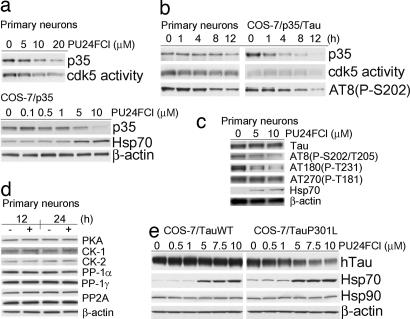
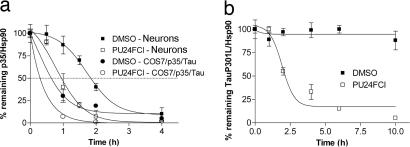
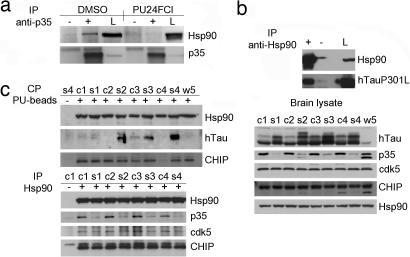
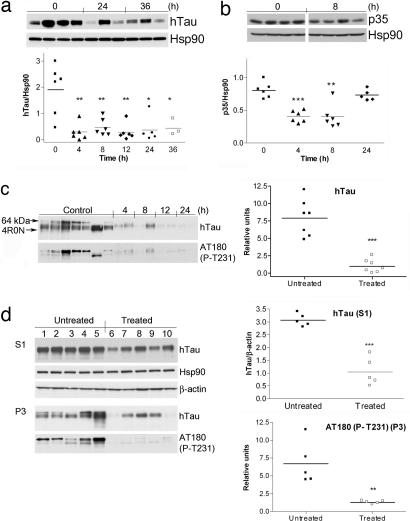
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
